INTRODUCTION
The genus Crepidiastrum Nakai belongs to the Asteraceae (Compositae) family, with approximately 15 species distributed from Central to Eastern Asia (Shih and Kilian, 2011). The genus Crepidiastrum has been divided into three sections; section Crepidiastrum, section Monostemma Nakai, and section Paraixeris (Nakai) Pak & Kawano, based on the habit, inflorescence position, number of inner phyllaries and florets, achenes shape (Pak and Kawano, 1992).
Among the taxa in the genus Crepidiastrum, C. ×muratagenii H. Ohashi & K. Ohashi, C. ×nakaii H. Ohashi & K. Ohashi, C. ×semiauriculatum N. Yamam. & H. Ikeda, C. ×surugense (Hisauti) Yonek. have been reported as hybrid taxa (Ohashi and Ohashi, 2007; Yamamoto et al., 2009). Such hybrid taxa make it difficult to understand the taxonomic boundaries among species within the three sections.
Previous studies on Crepidiastrum hybrid taxa have mainly been conducted on C. ×nakaii [= ×Crepidiastrixeris denticulato-platyphylla (Makino) Kitam. nom. inval.] based on morphological comparison, artificial hybrid examination, cytological and molecular analysis (Ono and Satô, 1935; Saito et al., 2003, 2006). However, the other hybrid taxa have not been studied.
A plant “Hong-do-go-deul-ppae-gi” has been considered C. ×nakaii [= ×Crepidiastrixeris denticulato-platyphylla (Makino) Kitam. nom. inval.] (Lee, 1969; Lee, 1996), a hybrid between C. denticulatum (in sect. Paraixeris) and C. platyphyllum (in sect. Monostemma). However, Jang and Choi (2021) suggested that the species should be considered C. ×muratagenii, a hybrid between C. denticulatum (in sect. Paraixeris) and C. lanceolatum (in sect. Crepidiastrum), based on its morphological traits and geographical distribution. However, Jang and Choi (2021) only examined individuals from Hongdo Island in based on analysis of nuclear ribosomal internal transcribed spacer regions (nrITS), and Hongdo Island individuals did not differ in sequence from C. denticulatum. Therefore, Hong-do-go-deul- ppae-gi has not yet been clearly identified taxonomically. Hybridization usually occurs in geographical locations where the parent species are sympatric. Hence, it is necessary to observe individuals in an area where C. denticulatum and C. lanceolatum co-occur. Therefore, Hongdo Island is unsuitable since only C. denticulatum is distributed in the region.
Molecular markers such as the nrITS regions have been used extensively to investigate hybrid speciation. Although in some cases, the utility of nrITS is limited in plant phylogenetic inference (Álvarez and Wendel, 2003), the molecular marker has been very useful in identifying hybrid taxa progenitors due to its biparental inheritance nature (Sang et al., 1995; Li, 2006; Du et al., 2009; Les et al., 2009; Høibová et al., 2011; Kokubugata et al., 2011). Since hybrid species originate by mixing genomes from two different species, detection of the parental genome in the putative hybrid taxa can be direct evidence of hybrid speciation. Furthermore, the addition of chloroplast DNA (cpDNA) data can help the detection and indicate the direction of hybridization in plants (Rieseberg et al., 1993; Schwarzbach and Rieseberg, 2002). Hence, it becomes necessary to examine not only internal transcribed spacer (ITS) but also cpDNA.
Additionally, previous studies (Kitamura, 1955; Saito et al., 2006; Ohashi and Ohashi, 2007) have suggested habit, margins of cauline leaves, number of involucral bracts and florets as identification keys of Crepidiastrum hybrids. However, such identification keys are not adequate, and additional morphological traits need to be reviewed.
In the present study, to reveal the hybrid origin of Hong-do-go-deul-ppae-gi, its morphological traits were observed based on specimens collected from other regions along with Hongdo Island. One nuclear ribosomal ITS region and four chloroplast regions (trnT-L, trnL-F, rpl16 intron, rps16 intron) of the putative hybrid and its closely related taxa were analyzed. Finally, the morphological traits were described.
MATERIALS AND METHODS
Morphological observation
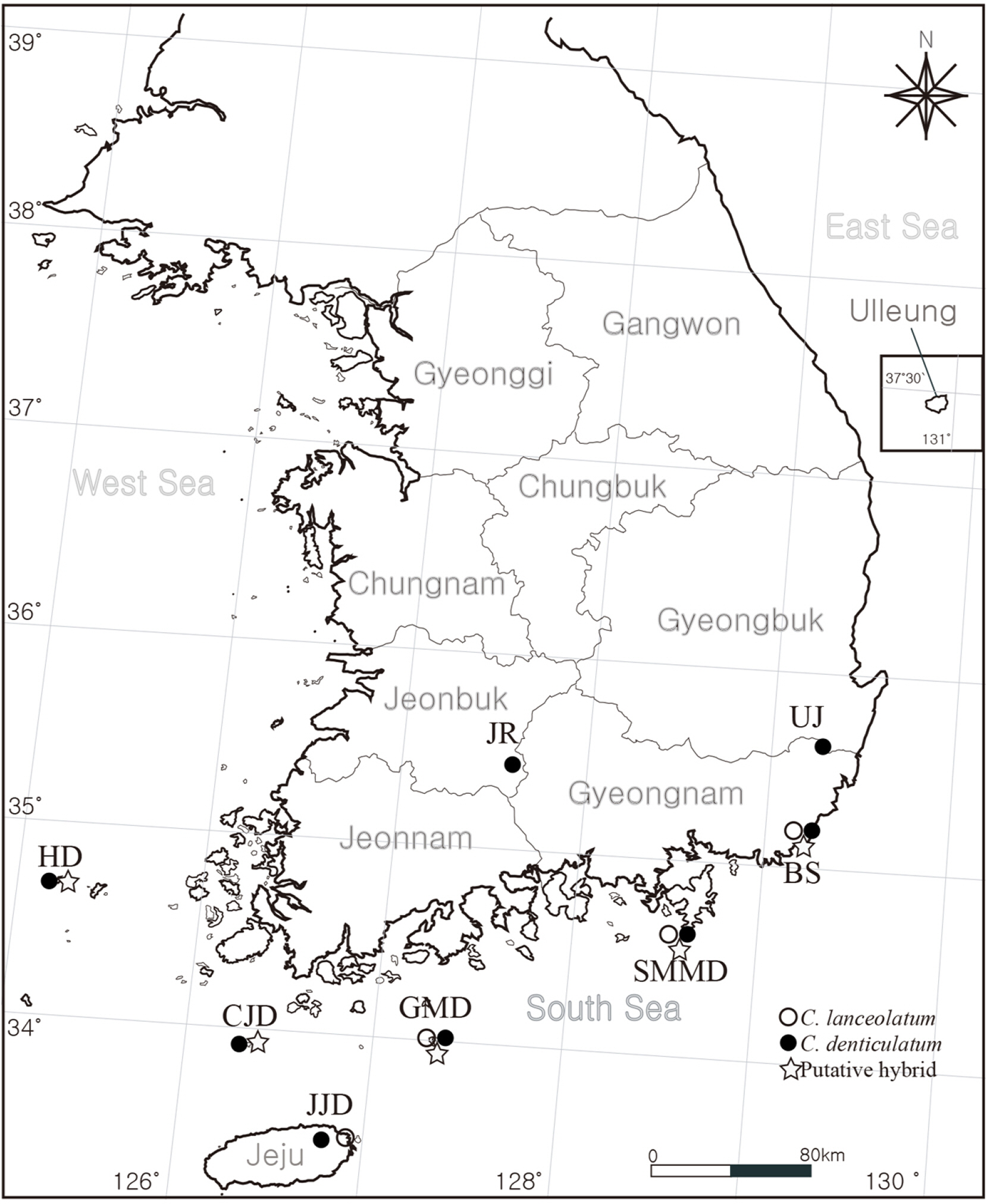
Twenty-four individuals were collected from seven localities in Korea (Fig. 1). To compare the morphological traits of the putative hybrid and its closely related taxa, we examined living materials and herbarium specimens stored at the herbarium of the Korea National Arboretum (KH), National Institute of Biological Resources (KB), Inha University (IUI) and registered at the Korea National Biospecies Information System (http://www.nature.go.kr), the collection database of specimens and materials (http://db.kahaku.go.jp/webmuseum_en/). Morphological traits were observed visually and under a stereomicroscope and sizes measured using a Mitutoyo 500-196-30 Absolute Digimatic Vernier caliper (Kanagawa, Japan). Additionally, protologues, flora, and monographs were referred to.
DNA extraction and PCR amplification
Leaf material from each individual was collected in silica gel for DNA extractions with voucher specimens (deposited in the herbarium of the Korea National Arboretum, KH). Total genomic DNA was extracted from dried leaf materials with silica gel using the DNeasy Plant Mini Kit (QIAGEN, Hilden, Germany) according to the manufacturer's instructions. The extracted DNA was electrophoresed in a 1% agarose gel to confirm the presence or absence of DNA. The concentration and quality of DNA were confirmed using a NanoDrop ND-1000 (NanoDrop Technologies, Wilmington, DE, USA).
Polymerase chain reaction (PCR) amplification of the ITS of the nuclear ribosomal DNA (nrDNA) and four noncoding regions (trnT-L, trnL-F, rpl16 intron, and rps16 intron) of the cpDNA was performed. The total volume of each PCR mix was 20 μL, comprising 15 μL of distilled water, 1.0 μL of each primer (50 mM), 1 unit of Taq DNA polymerase master mix (Amplicon, Rødovre, Denmark). The primers used for PCR amplification and the PCR cycle conditions are listed in Table 1. In addition, primers ITS 2 forward and ITS 3 reverse (White et al., 1990) were used as internal primers for sequencing confirmation in both directions, particularly in the hybrids that showed nucleotide polymorphisms. The PCR products were visualized in 1% agarose gels and sequenced using an ABI 3730xl DNA analyzer (Applied Biosystems, Foster City, CA, USA). The determined sequences were deposited in GenBank (Table 2). Additionally, C, platyphyllum individuals deposited in GenBank were used for phylogenetic analysis. The analyzed nucleotide sequences were determined after checking the chromatogram using Geneious R 7.1.9 (Biomatters Ltd., Auckland, New Zealand). All sequences were aligned using MUSCLE (Edgar, 2004). Polymorphic sites in the ITS region were identified by overlapping peaks on the chromatogram.
Phylogenetic analysis
Maximum likelihood trees based on the nrITS region and the combined four cpDNA regions (trnT-L, trnL-F, rpl16 intron, and rps16 intron) were constructed using the W-IQ-TREE Server (http://iqtree.cibiv.univie.ac.at/) (Trifinopoulos et al., 2016), based on user-friendly web servers for IQ-TREE 1.5 (Nguyen et al., 2015). Each aligned sequence dataset was tested to determine the best-fit model by using W-IQ-TREE with the Akaike criterion, and new model selection procedures. TIM3e was confirmed as the best-fit model for the selected nrITS and cpDNA regions. We evaluated the node supported by 1,000 ultrafast bootstrap replicates (UFBS) (Minh et al., 2013). Youngia japonica (L.) DC., which is most closely related to the genus Crepidiastrum, was used as an outgroup (Kilian et al., 2009).
RESULTS
Observation of morphological traits
As a result of observing the morphological traits, putative hybrid individuals were classified into three morphological types. All morphotypes had 10–12 florets but lacked basal leaves at anthesis. They grow on sunny slopes that were adjacent to mountains and coasts. However, Type 1 had herbaceous and erect stem branching in the upper part, mixed entire and serrate margins, 1.2–1.8 mm outer phyllaries, and achenes with 0.5–0.9 mm beaks. Type 2 had a suffrutescent stems with stout and short caudex, branched at the base, mixed entire and serrate margins, 1.4–1.6 mm outer phyllaries, and achenes with 0.2–0.5 mm beaks. Type 3 had a suffrutescent stem with stout and short caudex, branched at the base, mixed entire and serrate margins, 0.4–0.7 mm outer phyllaries, and achenes with 0.5–0.7 mm beaks. All three morphotypes showed that C. denticulatum and C. lanceolatum had mixed morphological traits (Fig. 2, Table 3). Type 1 and Type 2 were observed in Busan, Type 2 was observed on Somaemuldo Island, Type 2 and Type 3 were observed on Geomundo Island, and Type 3 was observed on Chujado and Hongdo Islands.
nrDNA ITS sequences and phylogenetic analysis
The sizes of the nrITS regions in C. platyphyllum, C. denticulatum, C. lanceolatum were 640 bp, 639 bp, and 638 bp, respectively. Size variation among the individuals within the same species was not observed. Four accessions of C. platyphyllum had identical sequences, while five accessions of C. denticulatum were grouped into three ribotypes (A, B, C) which differed at nucleotide position of 603, and five accessions of C. lanceolatum were grouped into three ribotypes (D, E, F) that differed at three nucleotide sites (200, 212, 434 bp) (Table 4). After alignment, a data set with a length of 641 bp was obtained. The length variation of the aligned sequence data was due to 1 to 3 bp of indels (insertion/deletion events). C. platyphyllum had 249 bp (1 indel) for the nrITS 1 region, 164 bp for the 5.8S, and 227 bp for nrITS 2 region. C. denticulatum and C. lanceolatum had 250 bp for the nrITS 1 region, 164 bp for 5.8S, but 225 bp (2 indels) and 224 bp (3 indels) for the nrITS 2 region, respectively. Excluding the gaps, the putative parental species showed sequence variation at a total of ten nucleotide sites. The pairwise-sequence difference between C. denticulatum and C lanceolatum was six nucleotide substitutions and one indel. C. denticulatum and C. platyphyllum differed by eight nucleotide substitutions and three indels. C lanceolatum and C. platyphyllum could be distinguished by six nucleotide substitutions and four indels. Therefore, the three taxa of Crepidiastrum were clearly distinguished in the nrITS regions. Among the putative hybrid types, Type 1 and Type 2 exhibited polymorphic ITS signals at those seven sites differentiating C. denticulatum and C. lanceolatum, showing sequence additivity. These were visible as double peaks in the electropherograms. Type 3 exhibited species-specific nucleotides of C. denticulatum. Additionally, no species-specific nucleotides from C. platyphyllum were observed in all putative hybrid individuals (Table 4).
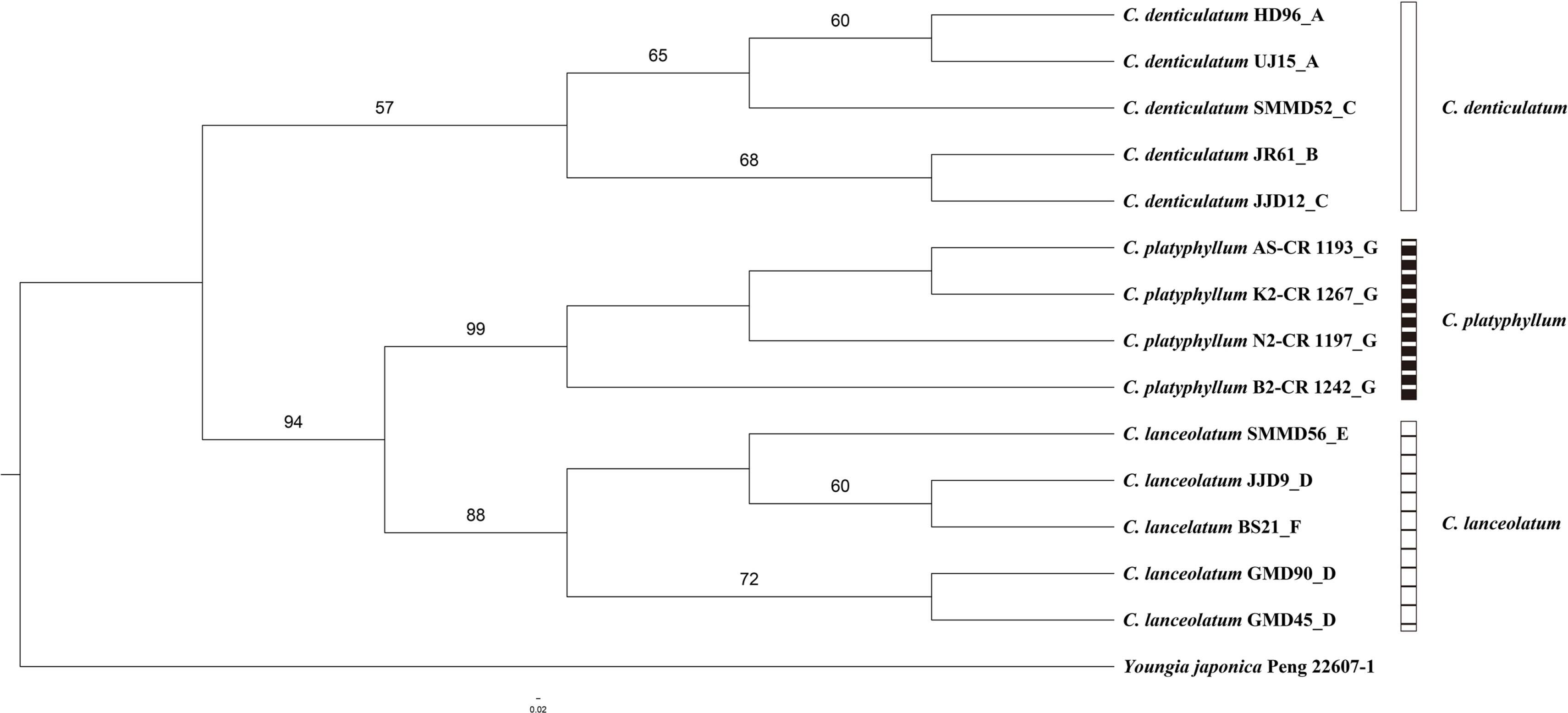
The aligned nrITS sequences of the three Crepidiastrum taxa except for the hybrid putative individuals had a length of 648 characters, comprising 546 constant, 90 parsimony-uninformative characters, and 12 parsimony-informative characters. In the nrITS phylogenetic tree, three taxa of the genus Crepidiastrum were divided into two clades. C. denticulatum was formed in a weakly supported clade (bootstrap [BS] = 57). Crepidiastrum platyphyllum and C. lanceolatum were formed in a clade (BS = 94), and each taxon was formed in a monophyletic group, respectively (BS = 99, BS = 88) (Fig. 3).
Combined cpDNA sequences and phylogenetic analysis
To trace the plastid donors (maternal origin) of the putative hybrid individuals, DNA sequences of the four plastid markers (trnT-L, trnL-F, rpl16 intron, and rps16 intron) were examined for the 24 individuals representing the putative hybrid and its closely related taxa. The size ranges of the four markers observed from the examined taxa were 528 to 537 bp (trnT-L), 766 bp (trnL-F), 949 bp (rpl16 intron), 721 to 740 bp (rps16 intron), respectively. In the trnT-L, sequence length for all examined accessions was 536–537 bp except C. denticulatum, C. platyphyllum, and Type 1 which had a length of 528–530 bp due to an 8 bp deletion at nucleotide sites 4 to 11, 1 bp insertion at nucleotide site 217 and single nucleotide variation observed at nucleotide position of 18. In trnL-F, two haplotypes (A, B) were found in C. denticulatum that showed single nucleotide variation at nucleotide position of 632. In rpl16 intron, three nucleotide variations were observed at nucleotide positions of 132, 307, and 801, respectively. The rps16 intron showed two nucleotide variations at nucleotide positions of 438 and 697 and two haplotypes were found in C. lanceolatum, which differed in size (721 and 740 bp). Therefore, among the analyzed four cpDNA markers, trnT-L, rpl16 intron and rps16 intron are useful for distinguishing between C. denticulatum and C. lanceolatum. However, since C. denticulatum and C. platyphyllum differ by only a single nucleotide sequence in the rpl16 intron (at 307 bp), additional markers are required to clearly distinguish the two taxa. A plastid type of C. platyphyllum was not detected in the examined samples of all the hybrid types (Table 5).
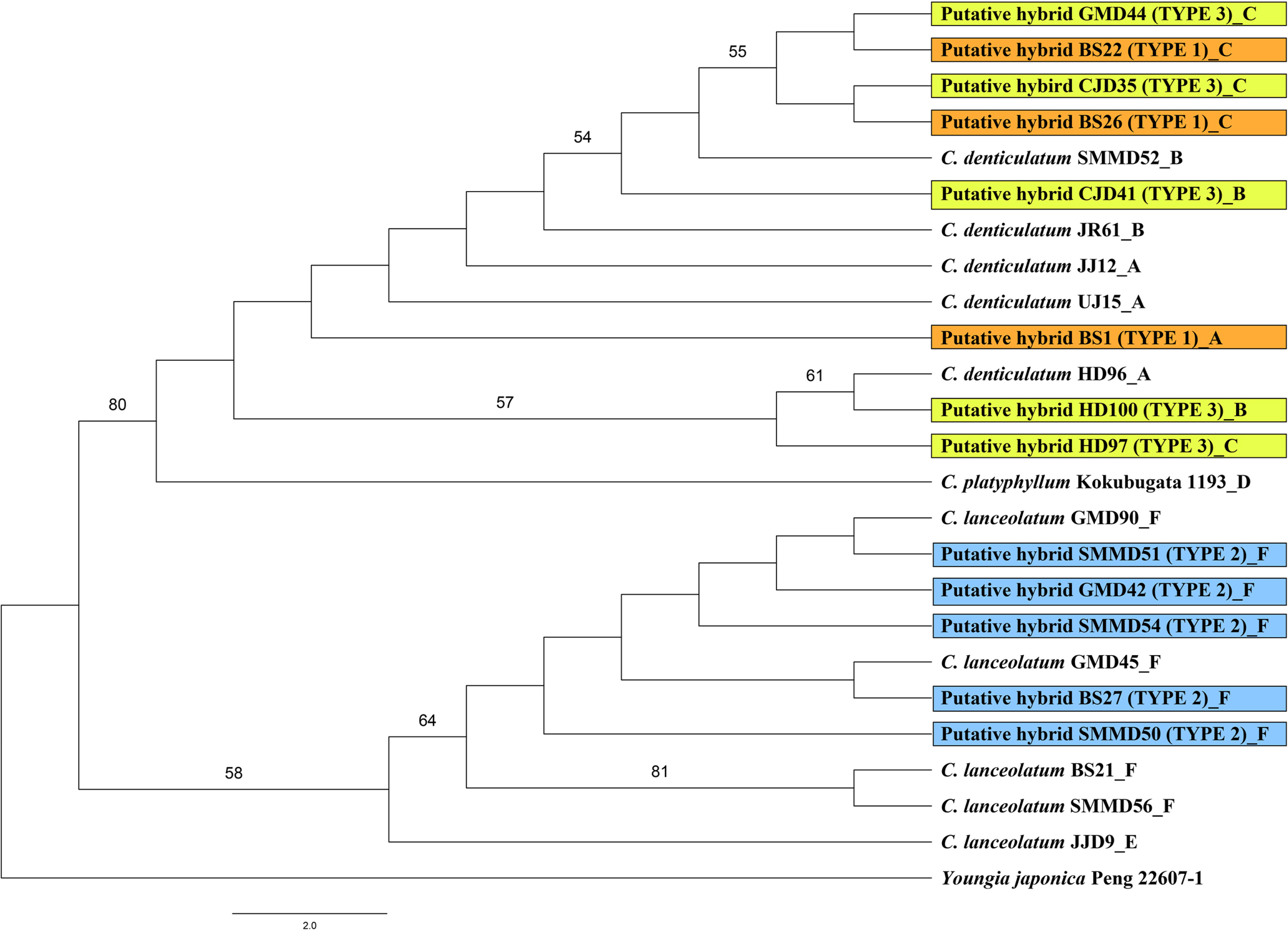
The aligned combined chloroplast sequence (trnT-L, trnL-F, rpl16 intron, and rps16 intron) had a length of 3,036 characters, comprising 2,959 constant, 69 parsimony-uninformative characters, and 8 parsimony-informative characters. In the combined cpDNA phylogenetic tree, the putative hybrid individuals and the three taxa of the genus Crepidiastrum were divided into two clades. Crepidiastrum denticulatum, C. platyphyllum, Type 1, and Type 3 formed a strongly supported clade (BS = 80%), and C. lanceolatum and Type 2 a weakly supported clade (BS = 56%) (Fig. 4).
DISCUSSION
The plant “Hong-do-go-deul-ppae-gi” has been regarded as C. ×nakaii, a hybrid between C. denticulatum and C. platyphyllum (Lee, 1969; Lee, 1996). However, the results of the molecular analysis of nrITS sequences of Type 1 and Type 2 individuals in the present study showed additive species-specific sites for C. denticulatum and C. lanceolatum (Table 4). This is interpreted from the results obtained by amplification of copies inherited from both parental species. Therefore, the results of the present study support the hypothesis proposed by Jang and Choi (2021), that Hong-do-go-deul-ppae-gi should be considered as C. ×muratagenii, a hybrid between C. denticulatum and C. lanceolatum. Nevertheless, Jang and Choi (2021) treated Hongdo Island individuals as a C. ×muratagenii. Type 3 included Hongdo Island individuals which had habitats and morphological traits similar to those of other hybrid types, but was distinguished by minute outer phyllaries (ca. 0.5 mm vs. ca. 1.5 mm in Type 1 and Type 2) (Fig. 2), which was identical with the sequence of C. denticulatum when the molecular analysis was performed. Hence, it was judged to be an ecological variation of C. denticulatum. Species-specific markers of C. platyphyllum were not detected in any putative hybrid individuals (Table 4). This agrees with the morphological observations, indicating that C. denticulatum and C. lanceolatum co-occurred in Busan, Somaemuldo and Geomundo Islands, and were involved in the hybridization.
The combined cpDNA sequences (trnT-L, trnL-F, rpl16 intron, and rps16 intron) of Type 1 and Type 2 showed species-specific markers for C. denticulatum and C. lanceolatum, respectively (Table 5). The results indicate that both C. denticulatum and C. lanceolatum have served as a plastid donor suggesting that, in the case of C.×muratagenii, the hybrid speciation that has occurred is bidirectional. However, no apparent geographic structuring of the haplotypes could be found. Further sampling might reveal the geographic structuring of the genus Crepidiastrum. In the combined cpDNA phylogenetic tree, C. denticulatum in sect. Paraixeris and C. platyphyllum in sect. Monostemma formed a clade (Fig. 4). However, the ITS phylogenetic tree showed that C. platyphyllum is sister to C. lanceolatum in sect. Crepidiastrum (Fig. 3). Similarly, additional sampling and markers are required to understand the relationships among the three sections and the relationships between species within a particular section.
In general, divergent ribotypes in hybrid lineages can be homogenized quickly by a mechanism known as concerted evolution (Wendel et al., 1995; Aguilar et al., 1999). However, some early generations of hybrid lineages can maintain divergent ribotypes contributed by parental species (Siripun and Schilling, 2005; Liu et al., 2009; Cho et al., 2014; Shin et al., 2014; Gil and Kim, 2016). The high number of completely additive polymorphic sites and the lack of new unique mutations in C. ×muratagenii in relation to its completely differentiated and non-sibling parental species are consistent with the expected pattern of recent or F1 hybrid formation (Rieseberg et al., 1993; Nieto Feliner et al., 2004).
The possibility of a backcross from one of the putative parents cannot be ruled out. However, we have not observed any such cases in the present molecular data, or perhaps our materials are too limited for drawing such conclusions. In other cases, regarding the C. ×nakaii from Saito et al. (2006), second or more generations have been reported and assumed. Also, the hybrid individuals observed in field research show a certain degree of fertility. There also exist a high likelihoods of backcrosses and introgressions.
The ITS and combined cpDNA (trnT-trnL, trnL-trnF, rpl16 intron, and rps16 intron) investigations provide strong evidence for the natural hybridization between C. denticulatum and C. lanceolatum. Further detailed investigations need to be performed on the pollen and ovule sterility, chromosome number, karyotype, and reproductive behavior of the hybrids.
Taxonomic treatment
Crepidiastrum ×muratagenii H. Ohashi & K. Ohashi, J. Jap. Bot. 82: 342, 2007.—TYPE: JAPAN. Kyushu: Kagoshima Pref., Yakushima, Yaku-cho, Kurio, 10 Nov 1983, G. Murata, T. Takagi & A. Iwami 40 (holotype: KYO, photo!, isotype: KYO, photo!).
= Crepidiastrum denticulatum (Houtt.) J. H. Pak & Kawano × C. lanceolatum (Houtt.) Nakai.
[×Crepidiastrixeris denticulato-lanceolata Kitam., Acta Phytotax. Geobot. 11: 132, 1942 nom.inval.]
Korean name: Hong-do-go-deul-ppae-gi (홍도고들빼기).
Herbs with erect stem and branched in the upper part or subshrubs with woody caudex stout, short, branched in the lower part, 10–30 cm tall. Leaves alternate, sessile; basal leaves withered at anthesis; blade of lower and middle cauline leaves spatulate-oblong, ovate-oblong or ovate, 4.1–7.9 × 1.5–4.2 cm, apex acute or obtuse, clasping at base, margins mixed entire and acute serrations, slightly thick texture; upper cauline leaves similar to middle leaves, 1.6–3.6 × 0.5–1.6 cm. Heads ca. 1.5 cm diam., with 10–12 florets; peduncle slender, 6.1–8.5 mm long; bracteoles many, ovate, 0.5–0.7 mm long; involucre tubular 6–8 mm long, with small bracteoles at base; outer phyllaries few, ovate, 1.2–1.8 mm long, acute or obtuse; inner phyllaries, 8, lanceolate, 5.5–7.4 mm long. Achenes fusiform, slightly compressed, brownish, 2.9–3.9 mm long, with 0.2–0.9 mm beak, 10–15 scabrid ribs; pappus 3–4 mm long, white.
Phenology: Flowering October to November and fruiting November to December
Distribution: China, Japan (Kyushu), Korea (Jeollanam-do, Gyeongsangnam-do).
Habitat: Sunny slopes adjacent to the mountains and the coast where C. denticulatum and C. lanceolatum occurred sympatrically.
Taxonomic notes: It is difficult to distinguish C. ×muratagenii because it shows mixed traits of C. denticulatum and C. lanceolatum (Fig. 2, Table 3). Crepidiastrum ×muratagenii was similar to C. lanceolatum by having outer phyllaries of 1.2–1.8 mm long (ca. 0.5 mm long in C. denticulatum), 10–12 florets (13–15 in C. denticulatum). However, it differed from C. lanceolatum by lacking basal leaves at anthesis, cauline leaves margins mixed with entire and acute serrations.