물통이속(Pilea Lindl.)은 쐐기풀과(Urticaceae)의 가장 큰 속으로, 전 세계적으로 600−700분류군이 속한다고 알려져 있으며(Adams, 1970; Burger, 1977; Monro, 2004), 암꽃의 화피는 3−5개의 열편으로 갈라지고, 주두는 붓 형태의 특이한 형태를 이루며, 수술이 튀어나왔을 때 꽃가루가 바람이 불어서 흩어지듯 동시에 흩어지는 특징을 가져 대포식물(artillery plant)이라고도 한다. 본 속의 식물들은 오스트레일리아와 뉴질랜드를 제외한 열대, 아열대 및 온대 지역에 넓게 분포하고 있으며, 속 내 대부분의 분류군은 육질성이고, 그늘이 있는 서늘한 지역을 좋아하며, 초본성, 아관목성, 관목성 등 다양한 식물형(plant habit)을 지닌다(Heywood, 1991).
본 속의 분류군에 대한 분류학적 연구는 Weddell (1856, 1869)이 엽형과 엽연의 형태로 Integrifoliae Weddell, Heterophyllae Weddell 와 Dentatae Weddell 의 3절로 구분하였던 것이 처음이다. Chen (1982, 1995)은 암꽃 화판의 수, 암꽃 화판의 형태, 엽맥의 형태, 수꽃 화서의 구조 등을 분류형질로 적용하여, Achudemia (Blume) C. J. Chen, Smithiella Dunn ex C. J. Chen, Tetrameris C. J. Chen, Pilea, Dimeris C. J. Chen, Urticella Miquel in Martius, Lecanthoides C. J. Chen 등의 7절로 구분한 바 있으며, Chen and Monro (2003)에 의해 속의 분류학적 연구가 7절로 최종적으로 수정되었다(Table 1).
우리나라에는 5종의 물통이속 식물이 자생하고 있으며 환경에 대한 적응력이 높아 어느 지역이든 쉽게 적응하여 활착하지만 특히, 그늘이 지고 습한 지역에 주로 분포하고 있으며, 해발고도가 낮은 지역에는 P. peploides (Gaudich.) Hook. & Arn. (물통이), P. mongolica Wedd. (모시물통이), P. hamaoi Makino (큰물통이) 등이 자생하고, 해발 고도가 높은 지역에는 P. taquetii Nakai (제주큰물통이), P. japonica (Maxim.) Hand.-Mazz. (산물통이)가 분포하고 있다. 종간에서는 자생하는 환경은 습한 지역으로 비슷하지만, 지리적인 격리와, 해발의 차이로 인해 형태적인 차이가 발생하고 있다.
선행되어진 한국산 물통이속에 대한 연구로는 종의 화학적 성분에 관한 연구(Kwon, et al., 1997; Ahn, 2003)가 전부로 자생하는 물통이속 식물에 대한 분류학적 연구는 찾기 힘들고, 분자계통학적인 연구는 전무한 상황이다. Moon (2011)의 석사학위 연구에서는 물통이와 제주큰물통이, 산물통이는 식물체의 크기, 잎의 형태, 화서의 형태에서 쉽게 구분되며, 모시물통이와 큰물통이는 완전히 성숙하기 전에는 육안으로 확인하여 분류하기에 어려움이 있지만 완전히 성숙한 화서를 가지고 있을 때는 쉽게 분류가 가능하다고 발표되었다.
nrDNA인 ITS (internal transcribed spacer) 지역과 cpDNA인 psbA-trnH 지역은 식물의 계통분석에 널리 사용되고 있으며, 진화속도가 빠르므로 유전자간의 변이성이 높아 종간이나 속간의 유연관계와 진화 경향성을 파악하는 데 널리 사용되고 있다(Baldwin, 1992; Soltis et al., 1992).
우리나라에 분포하는 물통이(Pilea)속 5분류군은 7절로 구분한 Chen(1982)의 분류체계를 기준으로, 산물통이(P. japonica (Maxim.) Hand.-Mazz.)는 Achudemia절에 속하고, 물통이(P. peploides (Gaudich.) Hook. & Arn.)는 Dimeris절에 속하고, 나머지 큰물통이(P. hamoi Makino), 모시물통이(P. mongolica Wedd.), 그리고 제주큰물통이(P. taquetii Nakai)는 Urticella절에 속한다. 이들을 구분하는 주요한 형질은 화서, 잎의 가장자리, 엽병의 길이, 암꽃의 화피의 개수, 열매의 반점 등으로, P. hamoi, P. mongolica, 그리고 P. taquetii는 화서와 잎의 가장자리의 형태 등으로 분류하기에는 많은 어려움이 있다. 특히 P. hamoi와 P. mongolica는 화서가 완전히 성숙하기 전에는 그 크기나 모양이 서로 비슷하여 동정의 어려움이 있다. 특히 종을 구분하는 주요 형질인 잎의 형질은 생육환경의 변화에 대한 적응으로 변이가 심하여 동정에 큰 어려움을 주었다. 그리고 제주특산종인 P. taquetii 역시 다른 종과의 형태학적 형질의 분명한 차이를 확인하기 어렵다. 이 연구는 우리나라에 서식하는 물통이속 5종에 대한 분류학적 문제를 검토하여, 속 내 종간의 유연관계를 확인하고자 수행하였다.
재료 및 방법
재료
2009년 8월부터 2010년 11월까지 전국 각 지역에서 채집하거나 국립수목원의 표본관 (KH), 난대림연구소의 표본관(WTFRC)에 소장되어 있는 석엽표본을 사용하였다(Table 2). 직접 채집한 식물들은 석엽표본으로 제작하고, 일부는 70% 에탄올에 고정하여 액침표본으로 제작한 후 공주대학교 표본관(KNH)에 보관하였다.
DNA 추출
재료는 채집지마다 각 분류군을 1종씩 선정하여 일정 부위의 잎을 수집하여 각 개체 별로 실리카겔 봉투에 넣은 후, 완전히 건조시켰다. 건조시킨 재료는 분쇄기(retsch, MM200)로 파쇄한 후, DNA 추출 kit (DNeasy Plant minikit, Qiagen)를 사용하여 Total genomic DNA를 추출하였다.
PCR 및 염기서열 결정
각 DNA 마커를 50 μL의 중합효소연쇄반응(PCR)에 의해 증폭하였다. 반응 구성물은 5 μL의 10x Pfu DNA buffer, 1 μL의 10 mM dNTP, 양방향의 1 μL의 10 pmol 프라이머, 1 μL의 template DNA, 0.5 μL의 Pfu DNA 중합효소와 나머지는 증류수로 채워 50 μL를 정량하였다. ITS nrDNA 마커의 중폭을 위하여 ITS1 (5'-TCC GTA GGT GAA CCT GCG G-3')과 ITS4 (5'-TCC TCC GCT TAT TGA TAT GC-3')(White et al., 1990)를 사용하였다. 95℃에서 2분간 초기 해리반응을 하고, 95℃ 20초, 50℃ 40초, 72℃ 1분을 하나의 cycle로 하여 총 30cycle을 수행하고, 72℃에서 5분간 고정하였다. psbA-trnH 부위는 trnH (5' -CGC GCA TGG TGG ATT CAC AAT CC-3')와 psbA (5'-GTT ATG CAT GAA CAT AAT GCT C-3')를 사용하고 (Shaw et al. 2005) 80℃에서 5분간 초기 해리반응을 했다. 94℃에서 30초, 50−56℃에서 30초, 72℃에서 1분을 하나의 cycle로 하여 총 35cycle을 수행하고, 72℃에서 10분간 고정하였다. PCR product는 시판되는 Kit (Solgent, PCR purification Kit)를 이용하여 증폭된 DNA만을 정제하고, 1% agarose gel에 1 μL를 적재하여 농도를 확인하였다. 정제된 PCR 산물은 PCR 프라이머를 이용하여 양방향으로 염기서열을 결정하였다.
염기서열 정렬 및 계통수 제작
각 시료에 대한 정방향 및 역방향 염기서열은 Sequencher(ver4.8; Gene Codes Corporation, Ann Arbor, MI, USA)를 사용하여 조합(assembling)하였다. 조합된 염기서열은 MacClade 4.08 (Maddison and Maddison, 2000)를 거쳐 Clustal X (Thompson et al., 1997)를 사용하여 정렬하며, 다시 MacClade를 이용한 수작업으로 최종 정렬하였다. 정렬결과 발생한 gap은 결여형질(missing character)로 처리하였으며, 모든 형질은 동일한 가중치를 적용하였다. Parsimony analysis, Neighbor joining (NJ)는 PAUP ver.4.0b (Swofford, 2002)를 사용하여 수행하였다. Parsimony analysis는 Heuristic search를 이용하였으며, Heuristic search의 option으로 TBR branch-swapping algorithm을 적용하였다. 또한, Retention Index (RI: Farris, 1989)는 PAUP에서 구했다. 외군(outgroup)으로는 Boehmeria에 속하는 2종을 포함하였다(Table 2). 총 5종 30개의 시료와 외군 2종의 nrITS와 psbA-trnH 지역의 DNA 염기서열을 파시모니 및 Neighbor Joining (NJ) 방법으로 분석하여 Bootstrap consensus 계통수와 NJ 계통수를 각각 제작하여 논의에 활용하였다.
결과 및 고찰
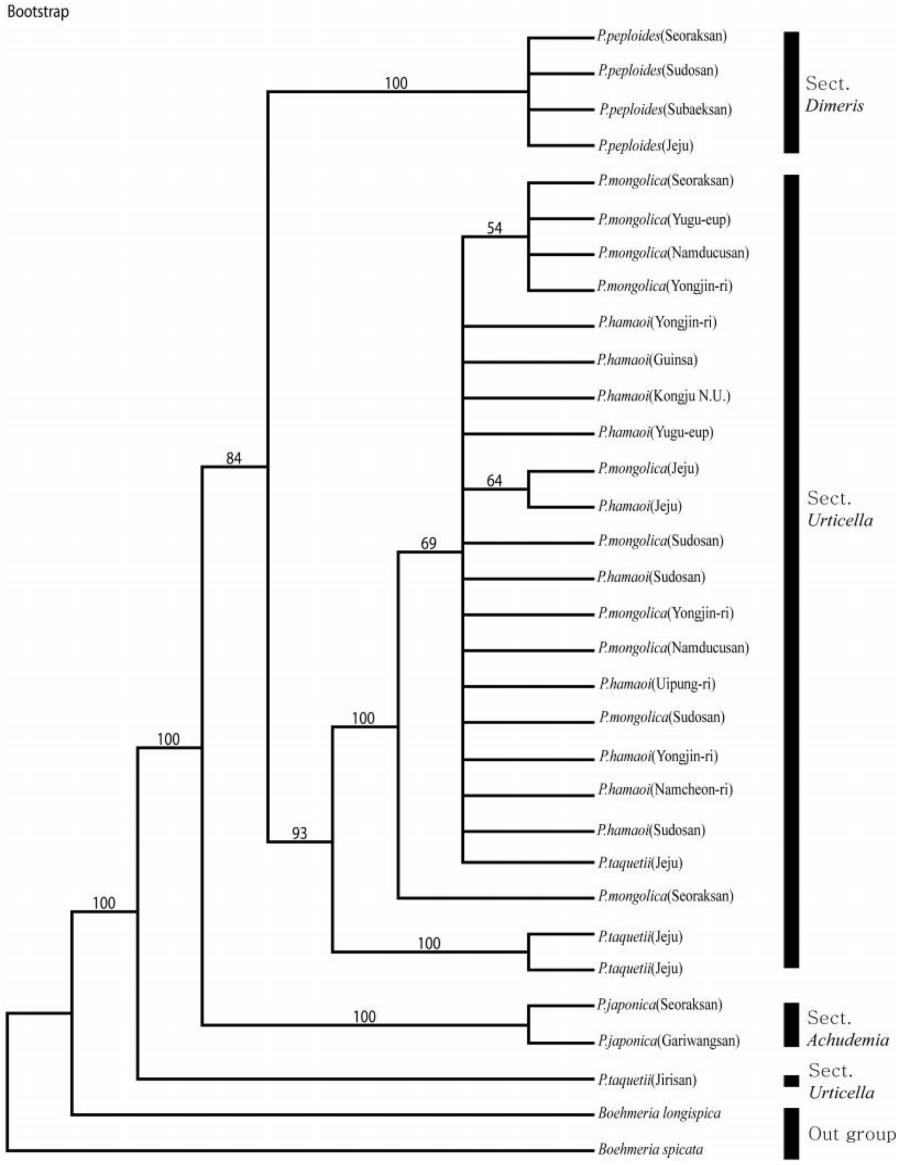
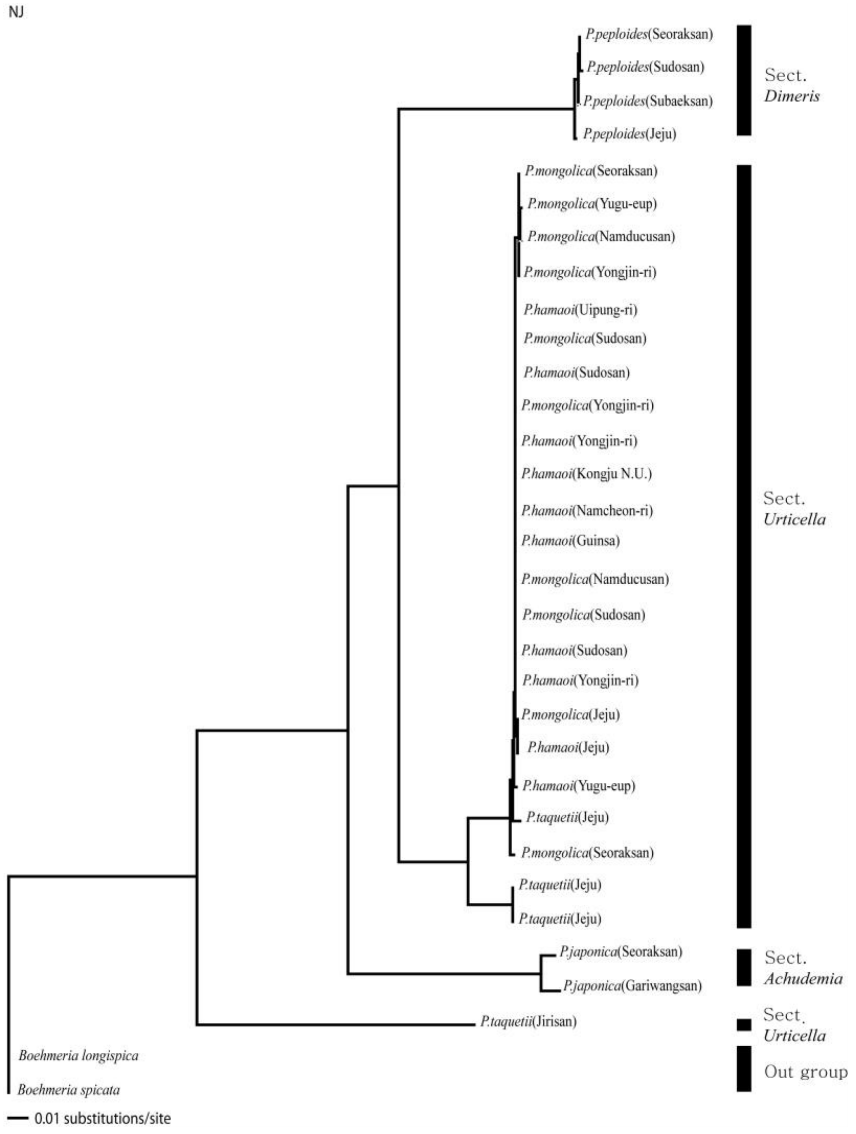
nrITS 지역의 DNA 염기서열의 길이는 517 bp로 정렬되었으며, 파시모니 분석 결과 일치계수 CI (Consistency Index)와 보유계수 RI (Retention Index) 값은 각각 0.905와 0.936로 높았으며, RC (Rescaled consistency Index) 역시 0.847로 비교적 높게 나타났다. 염기서열분석 결과 얻어진 NJ 계통수, Bootstrap 합의계통수는 Fig. 1−2과 같으며 이들은 모두 비슷한 양상으로 나타났다. 외군인 B. longispica와 B. spicata는 물통이속과 뚜렷이 분리되었다. P. taquetii는 화서가 성글게 생긴 것은 P. taquetii와 같지만 잎의 형태나 거치는 P. hamaoi와 유사성이 보이는 것이 하나의 분계조로 묶여 형성된 것으로 생각된다. 지리산 P. taquetii는 제주도의 P. taquetii와 하나의 분계조를 형성하지 않고 독립되어 나타나는데, 이 시료는 표본으로 확인하였으며, 화서는 성글게 맺히고, 잎도 연약한 것이 P. taquetii와 유사하지만 엽연이 확인된 다른 형질인 파상거치, 둔거치, 예거치, 치아상거치와는 다른 침상거치가 확인되었다. 이 결과, 지리산의 P. taquetii는 다른 분류군과 섞이지도 않아 다른 종으로 구분되어야 한다고 생각되며, 또한 중국에서 확인된 P. racemosa (Royle) Tuyama와 외부형태학적으로 엽형, 엽연, 화서의 길이 등이 유사한 것으로 보여 자세한 연구가 필요할 것으로 생각된다(Fig. 1). NJ tree의 분석결과, P. japonica가 다른 Pilea속 분류군과 분리되어 이들의 자매군으로 기부에서 분지하였으며, 나머지 P. hamaoi, P. mongolica, P. taquetii, P. peploides 중에서 P. peploides는 독립된 분계조를 형성하였다. 하나로 묶인 P. hamaoi, P. mongolica와 하나의 P. taquetii는 외부형태학적으로 잎의 형태와 화서 등의 구분이 모호한 것과 일치하는 것으로 생각되며, P. taquetii는 화서가 성글게 맺히는 것은 P. taquetii와 같지만 잎의 형태나 거치는 P. hamaoi와 유사성이 보이는 것이 하나의 분계조로 묶여 형성된 것으로, 이들의 유연관계가 높다고 생각된다. 지리산의 P. taquetii는 하나의 분계조로 형성되었는데, 이 시료는 표본으로 확인하였고, 직접 확인하지는 못했지만 화서는 성글게 맺히고, 잎도 연약한 것이 P. taquetii와 유사하지만 엽연이 확인된 다른 형질과는 다른 침상거치가 확인되었다. 이 결과, 지리산의 P. taquetii는 다른 분류군과 섞이지도 않아 다른 종으로 구분되어야 한다고 생각된다. 중국에서 확인된 P. racemosa (Royle) Tuyama와 외부 형태학적으로 유사성으로 보이는 것으로 생각되며, 이는 자세한 연구가 필요할 것으로 생각된다(Fig. 2).
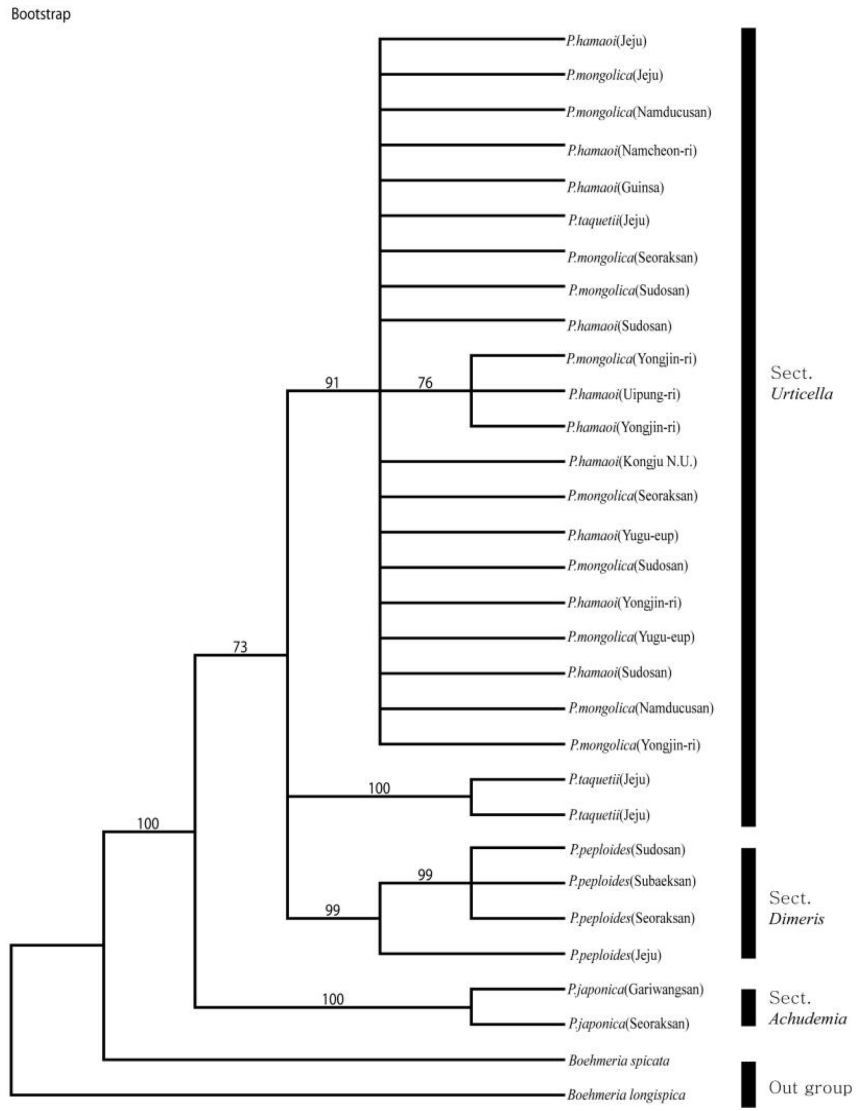
cpDNA의 psbA-trnH 지역의 염기서열 계통분석 결과, 일치계수 CI (Consistency Index)와 보유계수 RI (Retention Index)는 각각 0.843과 0.910였으며, RC (Rescaled Consistency Index)는 0.767이었다. 염기서열분석 결과 얻어진 NJ 계통수 및 Bootstrap 합의계통수는 Figs. 3−4와 같다. Bootstrap 계통수의 분석결과는 100%의 Bootstrap 값으로 P. japonica는 완전히 구분되어 분계조를 형성하였고, 다른 P. hamaoi, P. mongolica, P. taquetii, P. peploides는 73%의 Bootstrap 값으로 묶여 형성되었다. 이 분계조 안에서 P. peploides와 제주도의 P. taquetii 두 개체는 각각의 분계조를 형성하였다. 91%의 Bootstrap 값으로 묶인 P. hamaoi, P. mongolica와 하나의 P. taquetii는 외부형태적으로 잎의 형태와 화서 등의 구분이 모호한 것과 일치하는 것으로 생각되며, P. taquetii는 화서가 성글게 맺히는 것은 P. taquetii와 같지만 잎의 형태나 거치는 P. hamaoi와 유사성이 보이는 것이 하나의 분계조로 묶여 형성된 것으로 생각된다(Fig. 3).
NJ 분석결과 P. japonica는 완전히 구분되어 분계조를 형성하였고, 다른 P. hamaoi, P. mongolica, P. taquetii, P. peploides는 묶여 분계조를 형성했다. 이 분계조 안에서 P. peploides는 각각의 분계조를 형성하였다. 하나로 묶인 P. hamaoi, P. mongolica와 하나의 P. taquetii는 외부형태학적으로 잎의 형태와 화서 등의 구분이 모호한 것과 일치하는 것으로 생각되며, P. taquetii는 화서가 성글게 맺히는 것은 P. taquetii와 같지만 잎의 형태나 거치는 P. hamaoi와 유사성이 보이는 것이 하나의 분계조로 묶여 형성된 것으로, 이들의 유연관계가 높다고 생각된다(Fig. 4).
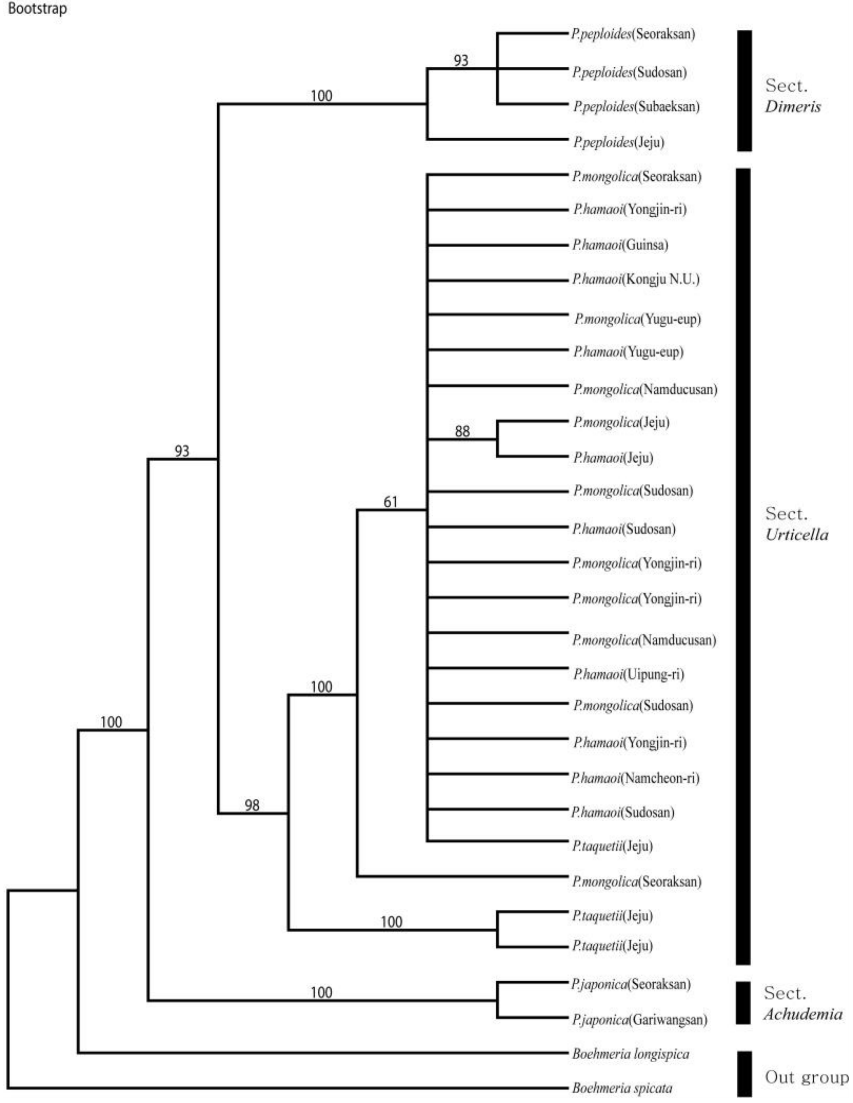
nrITS 지역과 cpDNA의 psbA-trnH 지역의 DNA 염기서열 자료를 통합하여 파시모니 분석을 실행한 결과 일치계수 CI (Consistency Index)와 보유계수 RI (Retention Index) 값은 각각 0.910과 0.952였으며, RC (Rescaled consistency Index)는 0.856이었다. 파시모니 분석 결과 얻어진 계통수는 Figs. 5−6과 같다.
NJ 계통수와 Bootstrap 합의계통수(파시모니 분석)는 모두 비슷한 양상으로 나타났다. 외군인 B. longispica과 B. spicata는 완전히 다른 분류군으로 기부에 분계조를 각각 형성하였다. Bootstrap 합의계통수의 분석결과는 100%의 Bootstrap 값으로 P. japonica는 완전히 구분되어 분계조를 형성하였고, 다른 P. hamaoi, P. mongolica, P. taquetii, P. peploides는 93%의 Bootstrap 값으로 묶여 형성되었다. 이 분계조 안에서 P. peploides는 100%의 Bootstrap 값으로 각각의 분계조를 형성하였으며, P. peploides는 내륙지역의 경남 수도산, 강원 설악산, 충북 소백산의 P. peploides가 93%의 Bootstrap 값으로 제주지역의 P. peploides와 구분되어 분계조를 형성하였는데, 이는 제주도에 자라고 있는 P. peploides가 지리적인 격리로 인해 내륙지역의 P. peploides와 약간의 차이를 형성한 것으로 생각된다. 61%의 Bootstrap 값으로 묶인 P. hamaoi, P. mongolica와 하나의 P. taquetii는 ITS1, 4 지역과 psbA-trnH 지역의 염기서열의 Bootstrap 합의계통수와 유사하게 결과가 형성되었으며, 외부형태학적으로 잎의 형태와 화서 등의 구분이 모호한 것과 일치하는 것으로 생각되며, P. taquetii는 화서가 성글게 맺히는 것은 P. taquetii와 같지만 잎의 형태나 거치는 P. hamaoi와 유사성이 보이는 것이 하나의 분계조로 묶여 형성된 것으로 생각된다(Fig. 5). NJ 계통수를 제작한 결과 P. japonica는 완전히 구분되어 분계조를 형성하였고, 다른 P. hamaoi, P. mongolica, P. taquetii, P. peploides는 묶여 분계조를 형성했다. 이 분계조 안에서 P. peploides와 P. taquetii는 각각의 분계조를 형성하였으며, 이는 내륙지역과 도서지역인 제주도의 지리적인 격리로 인한 계통학적으로 약간의 차이가 형성된 것으로 생각된다. 하나로 묶인 P. hamaoi, P. mongolica와 하나의 P. taquetii는 외부형태학적으로 잎의 형태와 화서 등의 구분이 모호한 것과 일치하는 것으로 생각되며, P. taquetii는 화서가 성글게 맺히는 것은 P. taquetii와 같지만 잎의 형태나 거치는 P. hamaoi와 유사성이 보이는 것이 하나의 분계조로 묶여 형성된 것으로, 이들의 유연관계가 높다고 생각된다(Fig. 6). 염기서열 분석에 사용된 시료 중 지리산에서 채집되었다는 P. taquetii의 표본에서 채취한 시료에서 nrDNA의 ITS 1, 4 지역의 염기서열 분석결과, Bootstrap 합의계통수 및 NJ 계통수에서의 공통적인 결과는 P. japonica와 제주지역의 P. taquetii, P. peploides는 각각의 분류군으로 분계조를 형성하지만 P. hamaoi와 P. mongolica는 clade내에서 산재되어 하나의 분계조를 형성하였다. P. peploides도 내륙지역과 제주지역의 P. peploides가 각각의 분계조를 형성하였다. 특히, 지리산의 P. taquetii는 외부형태학적으로 화서는 제주지역의 P. taquetii와 유사하지만, 분자계통학적 결과로는 한국산 물통이(Pilea)속에 속해 있는 다른 5분류군과의 유사성을 역시 보이지 않고 완전히 다른 종으로 구분되는 것으로 생각된다. 중국에서 확인된 P. racemosa (Royle) Tuyama와 외부 형태학적으로 유사한 것으로 생각되며, 국내의 미확인 종일 수도 있어서 차후에 자세한 연구가 필요할 것으로 생각된다.
본 연구의 nrDNA의 ITS 지역과 cpDNA의 psbA-trnH 지역 염기서열의 계통분석 결과에서는 한국에 자생하는 Pilea속의 5종은 Urticella, Dimeris, Achudemia 3절에 각각 속하고 있는 것으로 확인되었다. 다른 4종은 Chen (1982, 1995), Chen and Monro (2003)의 분류체계와 일치하고 형태적으로도 유사함을 결과적으로 확인하였지만 P. taquetii는 제주도에서 수집된 시료와 지리산에서 확인된 표본과의 유연성이 확인되지 않았다. 이는 도서지역과 내륙지역의 지리적인 격리로 인해 형태적인 변이가 생기고 이에 따라 시간이 흐르면서 유전적인 변화도 형성된 것으로 보인다. 지리산의 P. taquetii는 오히려 중국에서 확인된 P. racemosa (Royle) Tuyama와 잎, 화서 등의 형태가 유사하다는 것을 문헌으로 확인할 수 있었다. 이에 대해서는 표본으로만 확인한 지리산의 P. taquetii를 직접 확인하고 수집하여 중국의 P. racemosa (Royle) Tuyama와 제주도의 P. taquetii를 같이 연구하여 비교할 필요가 있을 것으로 생각된다.