쉬땅나무속(장미과)의 수리분류학적 연구
적 요
쉬땅나무속 분류군의 형태적 변이를 파악하고, 각 분류군의 식별형질을 검토하기 위해 정량적 형질을 이용하여 주성분분석(principal component analysis)을 수행하였다. 쉬땅나무속 분류군은 화서의 직립여부, 자방 및 골돌과 표면의 털 유무, 수술의 개수, 악편의 형태 등의 정성적 형질로 크게 두 그룹으로 구분된다. 본 연구 결과, 소엽의 개수 및 크기, 화서의 너비, 악편, 화판, 골돌과, 종자의 크기 등의 정량적 형질 또한 이들 두 그룹으로 뚜렷하게 구분되었다. 그룹 내에서는 Sorbifolia group (S. grandiflora, S. sorbifolia complex)의 경우, 측소엽의 크기와 엽맥의 수, 소엽 하면 내 선점 및 성상모의 개수, 화판, 골돌과의 크기 등의 형질이 유용한 식별형질로 파악되었으며, Kirilowii group (S. arborea complex, S. kirilowii, S. tomentosa complex) 내에서는 잎의 형질 중 정소엽과 측소엽의 크기 및 하면의 선점 및 성상모의 빈도 등이 분류군을 인식하는 데 유용한 것으로 파악되었다. 본 속의 수리분류학적 연구 결과는 추후 종 하 분류군을 포함한 쉬땅나무속 전체의 분류학적 재검토 및 분류학적 한계 설정에 매우 유용할 것으로 판단된다.
주요어: 쉬땅나무속, 장미과, 형태분석, 단변량분석, 주성분분석
Abstract
We conducted principal component analyses using the quantitative characteristics of the genus Sorbaria to investigate and explore morphological variation and diagnostic characteristics. The genus Sorbaria was divided into two groups based on erect or pendulous inflorescence, the existence of hairs on the ovary and follicle surfaces, the number of stamens, and the shape of the sepal. As a result of our investigation and of a morphometric analysis, these two groups could be also classified using quantitative characteristics, in this case the number of leaflets, the size of the leaflets, the width of the inflorescence, the size of the sepal, the petal, and the follicles and seeds. In the Sorbifolia group (S. grandiflora and S. sorbifolia complex), the size of lateral leaflets, number of veins, gland and stellate density on the abaxial surface of leaflets, and the petal and follicle size were found to be useful identification characteristics. The terminal and lateral leaflet size and the gland and stellate density on the abaxial surface of the leaflets were found to be characters of taxonomic value for the Kirilowii group (S. arborea complex, S. kirilowii, and S. tomentosa complex). The results of the numerical analysis conducted here can provide valuable information to those reconsidering and delimiting a taxonomic revision of the genus Sorbaria.
Keywords: Sorbaria, Rosaceae, morphometric analysis, univariate analysis, PCA (principal component analysis)
쉬땅나무속( Sorbaria (Ser.) A. Braun in Asch.)은 전통적인 장미과(Rosaceae Juss.) 분류체계에 따라 조팝나무아과(Spiraeoideae Arn.)내 쉬땅나무족(Sorbarieae Rydb.)에 속하는 분류군으로, 낙엽성 관목이며, 기수우상복엽이고, 탁엽이 있으며, 심피가 5개이고, 기부에서 유합하는 등의 특징으로 과 내 다른 분류군과 구별된다( Lingdi et al., 2003; Lee, 2007; Brouillet, 2015). 본 속은 주로 한국, 중국, 일본 등 동아시아와 아프가니스탄, 인도, 파키스탄을 비롯한 중앙아시아 및 러시아 동부에 분포하는 것으로 알려져 있으며( Purohit and Panigrahi, 1984; Ikeda, 2001; Gu and Alexander, 2003; Lee, 2007; Henrickson, 2015), 일부 분류군은 북미, 유럽 등지에 관상으로 도입되어 재배되는 것으로 보고되어 있다( Bailey, 1949; Ball, 1968; Tomaszewski, 2001; Henrickson, 2015). 쉬땅나무속은 기준표본인 Spiraea sorbifolia L. ( Linnaeus, 1753)을 기반으로 Spiraea L. sect. Sorbaria Ser. ( Seringe, 1825)인 조팝나무속 내 하나의 절(section)로 인식되었고, 이후 Braun (1860)에 의해 속으로 승격되었으며, 기준종은 Sorbaria sorbifolia (L.) A. Braun in Asch.로 현재까지 인식되고 있다. 쉬땅나무속에 대한 첫 번째 종속지적 연구(monograph)로 Maximowicz (1879)는 본 속을 조팝나무과(Spiraeaceae (Juss.) Dumort.)의 Gillenieae Maxim.족에 포함시켜, 열매 성숙 시 화주의 위치, 꽃의 크기, 엽맥 등의 형질을 통해 총 4개의 종으로 인식하였다. 이후, Schneider (1905)는 잎의 형태적 특징을 통해 보다 세분화하여 7종 2변종으로 정리하였고, Rehder (1940)는 S. tomentosa (Lindl.) Rehder를 포함한 5종 2변종으로 인식하였다. 한편, Pojarkova (1971)는 처음으로 속 내 분류체계를 제시하며, 열매가 늘어지는 정도와 크기, 꽃의 크기로 Lindleyanae A. Pojark., Sorbifoliae A. Pojark., Pallasianae A. Pojark.인 세 개의 열(series)로 구분하여, 총 4종으로 정리하였다. 이후에도 쉬땅나무속은 지리적인 분포와 학자 간 종의 개념에 따라 2–10여 종으로 다양하게 인식되어 왔다( Willis, 1948; Bailey, 1949; Ball, 1968; Schönbeck-Temesy, 1969; Yü and Ku, 1974; Purohit and Panigrahi, 1984; Rahn, 1989; Mabberley, 1990; Ikeda, 2001; Tomaszewski, 2001; Gu and Alexander, 2003; Lee, 2007).
Rahn(1989)은 본 속의 분류학적 재고를 위해 자생지 분포 및 기준표본 확인 등 표본조사를 수행하였다. 이에 아시아 전역에 분포하고 있는 쉬땅나무속의 형질과 지리적 분포를 검토하여, 골돌과의 크기와 털의 유무, 악편의 형태, 수술의 개수 등과 같은 외부형태적 형질을 바탕으로 다음의 4종 1변종으로 본 속 내 분류군을 인식하였다: S. sorbifolia (= S. sorbifolia var. stellipila Maxim., S. stellipila var. incerta C. K. Schneid.), S. grandiflora (Sweet) Maxim. (= S. rhoifolia Kom.), S. kirilowii (Regel & Tiling) Maxim. (= S. assurgens Vilm. & Bois, S. sorbifolia var. dubia C. K. Schneid., S. arborea C. K. Schneid. var. arborea, S. arborea var. glabrata Rehder in Sarg., S. arborea var. subtomentosa Rehder in Sarg.), S. tomentosa (= S. olgae Zinserl.), S. tomentosa var. angustifolia (Wenzig) Rahn (= S. aitchisonii Hemsl.) 하지만, 종 또는 종 하 분류군 간 정량적 형질의 통계분석 등 객관적인 형질 검토가 이루어지지 않았고, 한반도 남쪽에 분포하는 분류군이 제외되어 있다.
국내에는 쉬땅나무( S. sorbifolia var. stellipila) 1 분류군이 자생하는 것으로 보고되어 있으며( Lee, 2007), 최근 중국에 자생하는 S. kirilowii의 국내 분포를 확인하여 좀쉬땅나무로 발표한 바 있다( Song et al., 2010). 또한, 이 두 분류군 이외에 잎 하면 털의 빈도에 따라 쉬땅나무의 종 하 분류군으로 점쉬땅나무( S. sorbifolia var. glandulosa Nakai; 현재 비합법명으로 재고된 학명), 청쉬땅나무( S. sorbifolia f. incerta (C. K. Schneid.) Kitag.)를 인식하고 있는데, 이들 종하 분류군의 실체와 기본종과의 객관적인 형질 차이, 비합법명으로서 학명 사용의 문제 등 분류학적 문제와 한계설정이 모호한 실정이다( Lee, 2007). 따라서, 본 연구에서는 한국, 중국, 일본 등 동아시아와, 아프가니스탄, 인도, 파키스탄을 비롯한 중앙아시아 및 러시아 동부에 분포하는 세계산 쉬땅나무속 분류군을 대상으로 가능한 많은 형태학적 형질을 관찰 및 측정하고, 종 간 또는 종 하 분류군 간 형태 변이의 범주를 파악하며, 각 분류군의 유용한 식별형질을 검토하기 위해 형태 형질을 이용한 주성분분석(principal component analyses, PCA)을 수행하였다. 또한, 기존에 제시된 주요 정성적 식별형질을 재검토하고, 추가적인 정량적 식별형질을 탐색하여, 객관적 형질 차이를 기반으로 종 하 분류군의 실체를 파악하며, 분류군의 한계를 설정하는 데 그 의의가 있다.
재료 및 방법
국내 현지조사를 통해 채집된 식물체와 다음의 국내·외 표본관으로부터 대여한 표본을 바탕으로 연구하였다(acronyms according to Thiers, 2018). 국내: 강원대학교 표본관(KWNU), 경희대학교 식물표본관(KHUS), 국립수목원 산림생물표본관(KH), 서울대학교 농업생명과학대학 수우식물표본관(SNUA), 국외: Herbarium of the Arnold Arboretum (A), University of Copenhagen (C), Harvard University (GH), Royal Botanic Gardens Kew (K), Missouri Botanical Garden (MO), New England Botanical Club (NEBC), and Institute of Botany, Chinese Academy of Sciences (PE). 국내 현지조사를 통해 채집된 식물체는 석엽표본으로 제작하여 경희대학교 식물표본관(KHUS)에 확증표본으로 보관하였다. 형질 측정 및 관찰에 이용한 총 333점의 표본 중 형질 측정이 가능한 260개체를 형태분석에 사용하였다( S. arborea var. arborea: 20점, S. arborea var. glabrata: 14점, S. arborea var. subtomentosa: 24점, S. grandiflora [ S. rhoifolia 포함]: 8점, S. kirilowii: 58점, S. sorbifolia var. sorbifolia: 45점, S. sorbifolia var. glandulosa [현재 비합법명으로 재고]: 66점, S. sorbifolia var. stellipila: 55점, S. sorbifolia f. incerta: 4점, S. tomentosa var. tomentosa: 36점, S. tomentosa var. angustifolia: 3점). 분류군 식별을 위해서 일차적으로 원기재문을 참조하였고, 기존의 문헌과 자생 국가의 식물상( Maximowicz, 1879; Schneider, 1905; Pojarkova, 1971; Rahn, 1989; Ikeda, 2001; Gu and Alexander, 2003; Lee, 2007; Henrickson, 2015)에서 제시한 분류군의 특징과 분포를 바탕으로 동정 및 분류 후, 각 기관 별로 정량적 형질을 재검토 하였다. 본 연구에서는 Rahn (1989)에 의해 제시된 분류체계를 바탕으로 4개의 그룹( S. grandiflora [ S. rhoifolia 포함], S. sorbifolia complex, S. kirilowii complex [ S. arborea complex 포함], and S. tomentosa complex)으로 구분하여 각 그룹 간 형질 차이를 검토하였다. 또한 종 하 분류군의 정확한 정량적 형질 비교 및 분석을 위해 골돌과 표면 털의 유무, 악편의 형태, 수술의 수, 화서의 직립 여부 등의 주요 식별 형질과 각 분류군의 지리적 분포를 고려하여, 확실하게 구분되는 Sorbifolia group ( S. grandiflora, S. rhoifolia, and S. sorbifolia complex)과 Kirilowii group ( S. arborea complex, S. kirilowii, and S. tomentosa complex) 두 그룹으로 나누어 각 그룹 내 종 간 종 내 분류군의 형질 변이 및 차이를 분석하였다. 형태 분석을 위한 분석단위(operational taxonomic unit, 이하 OTU)는 기존 문헌에 나타나는 주요 형질과 지리적 분포를 중심으로 설정하였다. Sorbifolia group은 러시아 바이칼호 및 동시베리아 등지에 분포하는 S. grandiflora와 S. rhoifolia를 포함하여, 극동 러시아와 중국 북부 및 북미에 분포하는 S. sorbifolia, 중국 동북 3성(지린성, 랴오닝성, 헤이룽장성) 일부와 주로 한국과 일본 홋카이도에 분포하는 쉬땅나무( S. sorbifolia var. stellipila), 국내에만 분포하는 점쉬땅나무( S. sorbifolia var. glandulosa; 현재 비합법명으로 재고된 학명), 한국과 일본에 분포하는 청쉬땅나무( S. sorbifolia f. incerta), 총 6개의 OTUs로 구분하여 분석에 사용하였다. Kirilowii group은 인도, 파키스탄, 아프가니스탄 등지의 중앙아시아에 분포하는 S. tomentosa var. tomentosa, S. tomentosa var. angustifolia를 포함하여, 중국 내륙과 중국 남서부 티베트까지 분포하며, 고도 및 잎 하면 선점의 빈도 차이로 구분되는 S. arborea var. arborea, S. arborea var. glabrata, S. arborea var. subtomentosa와 중국 동북부와 최근 국내 분포가 확인된 좀쉬땅나무( S. kirilowii) 총 6개의 OTUs로 구분하여 분석에 사용하였다.
Sorbifolia group (Fig. 2)
형태 분석에 사용된 형질은 Sorbifolia group에서 총 37개의 정량형질을 선정하였고, 이는 영양형질 21개, 생식형질 16개에 해당한다( Table 1, Appendix 1). 영양형질은 소엽의 개수(C1), 잎의 길이(C2), 엽병의 길이(C3), 정소엽의 길이(C4), 정소엽의 너비(C5), 정소엽 길이/너비의 비(C6), 측소엽의 길이(C7), 측소엽의 너비(C8), 측소엽 길이/너비의 비(C9), 정소엽 소엽병의 길이(C10), 정소엽 엽맥의 수(C11). 측소엽 엽맥의 수(C12), 측소엽 엽연 거치의 길이(C13), 측소엽 엽연 거치 원측면 길이(C14), 측소엽 엽연 거치 근측면 길이(C15), 소엽 정단부 선모의 개수(C16), 소엽 중부 선모의 개수(C17), 소엽 기부 선모의 개수(C18), 소엽 정단부 성상모의 개수(C19), 소엽 중부 성상모의 개수(C20), 소엽 기부 성상모의 개수(C21)이며, 생식형질은 화서의 길이(C22), 화서의 너비(C23), 악편의 길이(C24), 악편의 너비(C25), 악편 열편의 길이/너비의 비(C26), 화판의 길이(C27), 화판의 너비(C28), 화판의 길이/너비의 비(C29), 꽃의 직경(C30), 화사의 길이(C31), 열매의 길이(C32), 열매의 너비(C33), 열매의 길이/너비의 비(C34), 종자의 길이(C35), 종자의 너비(C36), 종자의 길이/너비의 비(C37)에 해당한다.
Kirilowii group (Fig. 6)
형태 분석에 사용된 형질은 Kirilowii group에서 총 34개의 정량형질을 선정하였고, 이는 영양형질 17개, 생식형질 17개에 해당한다( Table 2, Appendix 2). 영양형질은 소엽의 개수(C1), 잎의 길이(C2), 엽병의 길이(C3), 정소엽의 길이(C4), 정소엽의 너비(C5), 정소엽 길이/너비의 비(C6), 측소엽의 길이(C7), 측소엽의 너비(C8), 측소엽 길이/너비의 비(C9), 정소엽 소엽병의 길이(C10), 정소엽 엽맥의 수(C11). 측소엽 엽맥의 수(C12), 측소엽 엽연 거치의 길이(C13), 측소엽 엽연 거치 원측면 길이(C14), 측소엽 엽연 거치 근측면 길이(C15), 소엽 중부 선모의 개수(C16), 소엽 중부 성상모의 개수(C17)이며, 생식형질은 화서의 길이(C18), 화서의 너비(C19), 2차 화서 사이 화경의 길이(C20), 악편의 길이(C21), 악편의 너비(C22), 악편 열편의 길이/너비의 비(C23), 화판의 길이(C24), 화판의 너비(C25), 화판의 길이/너비의 비(C26), 꽃의 직경(C27), 화사의 길이(C28), 열매의 길이(C29), 열매의 너비(C30), 열매의 길이/너비의 비(C31), 종자의 길이(C32), 종자의 너비(C33), 종자의 길이/너비의 비(C34)에 해당한다. 분석에 사용한 잎은 개화기나 결실기에 완전히 성숙한 잎(측소엽의 경우 가장 큰 잎)을 측정하였으며, 꽃의 형질은 화서의 정단부에 있는 것을 측정하였다. 성숙엽 하면의 털의 종류와 빈도의 객관적 정량화를 위해 주맥과 엽연의 중간 부분에 측맥을 포함한 2×2mm2 구간에 분포하는 털의 개수를 측정하였다.
각 형질은 디지털 캘리퍼스(Digital Calipers, CD-15CP, Mitutoyo, Kawasaki, Japan)를 이용하여 측정하였고, 선점, 성상모의 빈도를 확인하기 위해 해부 현미경(streomicroscope, SteREO Discovery. V8, Carl Zeiss Microscopy GmbH, Jena, Germany)을 사용하여 개수를 정량화하였다. 각 형질의 종간 변이를 확인하기 위해서 조사된 형질은 단변량분석(최소, 최대, 평균, 표준편차)을 수행하여 기재하였고( Appendices 1, 2), 동일종이 다른 지역에 분포하는 경우, 지역 간 차이를 확인하기 위해 분포 국가 별로 구분하여 오차막대(평균, 95% 신뢰구간)로 표현하였다. 측정값은 표준화한 후, PCA를 수행하였다(IBM Corp. Released 2012. IBM SPSS Statistics for Windows, version 21.0. Armonk, NY, USA).
결 과
Sorbaria속 내 분류군간의 정량적 형질 비교
최근 분류체계에서 Sorbaria속 내 종으로 인식되고 있는 분류군을 S. grandiflora (SG), S. sorbifolia complex (SS cmx), S. kirilowii complex (SK cmx; S. arborea complex 포함), S. tomentosa complex (ST cmx) 네 그룹에 대한 형질 비교에서 S. grandiflora가 나머지 그룹에 비해 대부분의 영양형질 및 화서의 정량적 형질에서 작게 나타나 구별되었고( Fig. 1A– I), 생식형질 중 악편, 화판, 꽃의 직경, 열매의 형질에서는 크게 나타나 형질의 중복 없이 나머지 그룹과 확연히 구별되었다( Fig. 1J– L). S. sorbifolia complex와 S. kirilowii complex는 대부분의 영양형질에서는 형질의 중복이 나타났지만( Fig. 1B– G), 화서의 형질을 제외한 악편, 화판, 열매, 종자의 생식형질에서 S. sorbifolia complex가 모두 크게 나타나 중복 없이 구별할 수 있었다( Fig. 1I– L).
Sorbifolia group (S. grandiflora, S. rhoifolia, and S. sorbifolia complex) 내 정량적 형질 비교
1. 영양형질-잎, 소엽
21개 영양형질에 대한 PCA 결과, 고유값이 1.00 이상인 6개의 요인 중에서 주성분 1, 2, 3이 전체분산의 63.2%를 설명하였다( Table 3). 주성분 1은 전체 분산의 28.9 %를 설명하며, 잎의 크기(C2, C3), 정소엽(C4, C5), 측소엽(C7, C8)의 크기, 측소엽 엽연 거치의 길이(C13, C14, C15)의 형질들과 비교적 높은 상관관계를 나타내었고( Table 3), 주성분 2는 전체 분산의 20.4 %를 설명하며, 정소엽(C11)과 측소엽(C12)의 엽맥 수, 소엽 하면의 선점 개수(C16, C17, C18)등에 관련된 형질이 높은 vector값을 나타냈다( Table 3). 주성분 3은 기여율이 13.9%로 비교적 낮게 나타났고, 소엽 하면의 성상모 개수(C19, C20, C21)가 높은 vector값으로 나타났다( Table 3). 기여율이 가장 높은 주성분 1과 2를 축으로 하여 각 개체들을 배열한 결과, S. grandiflora- S. rhoifolia가 주성분 1과 2에서 낮은 값을 가지면서 왼쪽, 하단에 위치하여 나머지 분류군 개체들과 구분되었다( Fig. 3A). 이는 S. grandiflora- S. rhoifolia이 다른 분류군에 비해 소엽의 크기가 작으며, 거치가 얕게 결각되어 있고, 소엽의 엽맥의 수가 적음을 의미한다. 또한, 주성분 2에 의해 높은 양의 값을 갖는 점쉬땅나무가 S. sorbifolia complex 내에서 확연히 구별되었는데, 이는 소엽의 엽맥 수가 다른 분류군에 비해 상대적으로 많으며, 소엽 하면 내 선점의 빈도가 높은 것을 의미한다( Fig. 3A). 주성분 2와 3에 의한 2차원 도해에서는 성상모의 빈도가 높은 쉬땅나무와 선점의 빈도가 높은 점쉬땅나무가 다른 분류군과 구별되었다( Fig. 3B).
2. 생식형질-화서, 꽃, 열매, 종자
16개 생식형질에 대한 PCA 결과, 5개의 요인 중 주성분 1과 2는 각각 전체 변이의 34.9%, 13.1%를 설명하였다( Table 4). 주성분 1은 주로 화판(C24, C25, C30), 악편(C27, C28)의 크기, 열매의 크기(C32, C33)의 형질들과 높은 상관관계를 나타내었고( Table 4), 주성분 2는 화서의 너비(C23), 종자의 너비(C36) 형질에서 높은 값으로 기여하였다( Table 4). 기여율이 높은 주성분 1과 2에 의해 배열된 2차원 도해에서 S. grandiflora- S. rhoifolia이 주성분 1에서 높은 값을 가지며, 나머지 분류군과 확연히 구분되었고, S. sorbifolia complex는 혼재되어 나타났다( Fig. 4). 단변량 분석을 통한 오차막대 도해에서는 잎 하면 선모의 빈도가 한국에 분포하는 쉬땅나무 분류군에서 중국과 일본의 분류군에 비해 높았고, 점쉬땅나무는 형질의 중첩없이 단위면적 당 많은 수의 차이로 쉽게 구분되었다( Fig. 5D– F). 성상모의 빈도 역시 다른 분류군과 중첩 없이 높았으며, 특히 국내에 분포하는 분류군이 중국과 일본의 분류군에 비해 빈도가 높게 나타나 뚜렷하게 구별되었다( Fig. 5G– I). 또한, S. grandiflora, S. rhoifolia가 화판, 악편, 꽃의 직경에서 다른 분류군과 형질의 중첩 없이 독립적으로 구분되었다( Fig. 5J– L).
Kirilowii group (S. arborea complex, S. kirilowii, and S. tomentosa complex) 내 정량적 형질 비교
1. 영양형질-잎, 소엽
21개 영양형질에 대한 PCA 결과, 주성분 1, 2, 3이 전체 분산의 70.3%를 설명하였다( Table 5). 각각 전체 변이의 39.2%, 19.9%, 11.2%를 설명하며, 주성분 1은 전체 분산의 39.2%를 설명하고, 잎(C2), 엽병(C3)의 길이, 정소엽(C4, C5), 측소엽(C7, C8)의 크기, 소엽병의 길이(C10), 측소엽 엽연 거치의 길이(C13, C15)의 형질들과 비교적 높은 상관관계를 나타내었으며( Table 5), 주성분 2는 전체 분산의 19.9 %를 설명하고, 소엽 측맥의 수(C11, C12), 소엽 하면의 털의 빈도(C16, C17)와 관련된 형질이 높은 vector값을 나타냈다( Table 5). 주성분 3은 기여율이 11.2 %로 매우 낮게 나타났다( Table 5). 기여율이 가장 높은 주성분 1과 2를 축으로 하여 2차원 도해에서 각 개체들을 배열한 결과, 전체적으로 연속성을 보이지만 크게 S. kirilowii, S. tomentosa var. tomentosa와 S. arborea var. subtomentosa, S. tomentosa var. angustifolia로 구분할 수 있었다( Fig. 7A). 주성분 2와 3의 경우 크게 ( S. tomentosa var. tomentosa, S. tomentosa var. angustifolia), ( S. arborea var. glabrata, S. kirilowii), ( S. arborea var. arborea, S. arborea var. subtomentosa) 세 개의 그룹으로 구분할 수 있었다( Fig. 7B).
2. 생식형질-화서, 꽃, 열매, 종자
17개 생식형질에 대한 PCA 결과, 6개의 요인 중 주성분 1, 2, 3이 전체분산의 52.2 %를 설명하였다( Table 6). 주성분 1은 전체 분산의 22.1%를 설명하며, 주로 악편(C21, C22), 화판(C24, C25), 꽃(C27)의 크기 등의 형질들과 높은 상관관계를 나타내었고( Table 6), 주성분 2는 화사의 길이(C28), 열매(C29, C30), 종자(C32, C33)의 크기 형질에서 높은 값으로 기여하였다( Table 6). 주성분 1과 2에 의한 2차원 도해에서 S. tomentosa var. tomentosa와 S. tomentosa var. angustifolia는 두 분류군 모두 주성분 1에서 양의 값을 나타내 오른쪽에 위치하였으나, 주성분 2에 의해 S. tomentosa var. tomentosa는 하단에, S. tomentosa var. angustifolia는 상단에 위치하여 구별되었다( Fig. 8A). 이는 S. tomentosa var. angustifolia의 열매, 종자가 기본종에 비해 비교적 큰 것을 의미한다. 주성분 1과 2, 주성분 2와 3에서 모두 S. kirilowii와 S. arborea complex는 혼재되어 나타났다( Fig. 8A, B). 단변량 분석을 통한 오차막대 도해에서는 대부분의 영양형질에서 S. arborea var. arborea, S. arborea var. subtomentosa가 중국과 국내에 분포하는 S. kirilowii와 중첩없이 구분되었으며, S. arborea var. glabrata의 경우 두 그룹의 형질 변이를 모두 포함하는 양상을 보였다( Fig. 9). 하지만 잎 하면 선점의 높은 빈도는 S. arborea var. arborea, S. arborea var. subtomentosa를 다른 분류군들과 확연히 분류할 수 있었으며( Fig. 9G), 성상모의 빈도는 S. arborea var. subtomentosa에서 커다란 빈도의 차이를 보였다( Fig. 9H). 또한 대부분의 형질에서 S. tomentosa var. tomentosa와 S. tomentosa var. angustifolia 두 분류군은 중첩 없이 독립적으로 구분되었다( Fig. 9). 또한, 생식형질로는 2차 화서 사이 화경의 길이의 형질로 S. arborea var. arborea, S. arborea var. subtomentosa와 S. kirilowii를 독립적으로 구분할 수 있었다( Fig. 9I). 하지만 S. arborea var. glabrata는 잎의 형질에서와 같이 두 그룹의 형질을 모두 포함하여 중첩되는 양상을 보였다( Fig. 9A, C, E, F).
고 찰
쉬땅나무속의 주요 식별형질
쉬땅나무속 내 종 간 식별형질로는 크게 골돌과의 크기와 털의 유무, 악편의 형태, 수술의 개수, 화서의 직립 여부 등으로 구별되며, 분류군의 수는 Maximowicz (1879)가 제안한 4종을 기본으로 하고 있다( Rahn, 1989). 정성적 형질뿐 아니라 영양형질과 생식형질의 정량적 형질 분석을 통해서도 러시아에 분포하는 S. grandiflora ( S. rhoifolia)가 다른 세 그룹과는 확연히 구별되었고, 중앙아시아에 분포하는 S. tomentosa complex를 포함한 나머지 그룹 간에서 일부 형질을 제외하고, 중첩이 나타나지 않아 식별형질로서 정량적 형질 또한 유용한 것으로 나타났다. 또한, 개화기에 화서의 직립여부, 자방의 털의 유무, 수술의 개수, 악편의 형태, 결실기 골돌과 털의 유무 등으로 S. sorbifolia complex와 S. kirilowii complex를 구분할 수 있는데, 본 연구에서 소엽의 개수, 측소엽의 길이, 화서의 너비, 악편, 화판, 골돌과, 종자의 크기의 정량적 형질을 통해서도 이들 두 그룹을 구별할 수 있는 식별형질로 사용될 수 있음을 확인하였다. 따라서, 정량적 형질을 통한 결과 역시 속 내 분류체계를 크게 4분류군으로 구분하고 정리한 Maximowicz (1879)와 Rahn (1989)의 분류체계를 지지하였다.
PCA에 의한 식별형질 검토
1. Sorbifolia group
Schneider (1905)는 자방의 털의 유무, 악편의 형태, 잎의 형태 등으로 S. sorbifolia, S. stellipila (Maxim.) C. K. Schneid. 2종과 종 하 분류군으로 구별하였으며, 이 후 Rahn (1989)은 S. sorbifolia 한 종으로 통합하여 인식하였다. 본 연구에서는 Sorbifolia group ( S. grandiflora, S. rhoifolia, and S. sorbifolia complex) 내 모든 분류군에서 개화기 자방에 발달한 털을 확인하였고, 결실기 골돌과에서도 동일한 형태의 털을 확인 할 수 있었다. 또한 악편 및 잎의 형태 역시 Sorbifolia group 내에서 S. grandiflora를 제외하고 길이와 형태의 차이가 나타나지 않아 Schneider (1905)이 제시한 S. sorbifolia, S. stellipila를 구분하는 자방의 털의 유무, 악편 및 잎의 형태는 식별형질로서 유용하지 않은 것으로 확인되었다. Sorbifolia group의 주요형질을 이용한 PCA 결과, 소엽의 크기와 엽맥 수, 소엽 하면의 선점 및 성상모의 개수 등의 형질이 유용한 식별형질로 확인되었다. 이 중 성상모의 개수, 빈도는 쉬땅나무( S. sorbifolia var. stellipila)에서 분포지역에 관계없이 높게 나타나, Maximowicz (1879)의 종하 분류군의 인식을 지지하였으며, 선점의 개수는 처음 선점의 형질로 분류군을 인식한 Nakai의 의견을 지지하였다. 하지만, 점쉬땅나무, S. stellipila var. glandulosa Nakai ex T. Kawamoto의 학명은 鮮滿實用林業便覽 ( Kaburaki, 1940)에 발표된 비합법명으로 라틴 기재문 없이 일본명, 학명, 일본어로 종의 실체를 언급하고 있어( Chang and Kim, 2002; Kim et al., 2005), 국제식물명명규약(International Code of Nomenclature, ICN) ( McNeill et al., 2011), Melbourne Code 39.1, 41.5에 의거하여 비합법명이 된다. 또한, 이러한 나명(Nomen nudum)의 기본명을 조합하여 처리한 Lee (1980)의 S. sorbifolia f. glandulosa (Nakai) T. B. Lee의 학명 역시 사용할 수 없다. 본 연구 결과뿐 아니라, 잎의 해부학적( Song and Hong, 2014), 미세형태학적 연구( Song and Hong, 2016a)에서도 잎 하면에 높은 빈도로 존재하는 선점의 형질로 점쉬땅나무를 다른 분류군과 구분하고 있다. 본 연구에서는 점쉬땅나무의 분포가 국내 강원도에 한정되어 있고, 선점의 빈도가 확연히 구분되며, 성상모와 선점이 함께 분포하는 개체에서도 그 빈도 차이가 확실히 구분되어, 변종 수준의 종하 분류군으로 인식하는 것이 바람직하다고 판단된다. 이에 모든 형질의 종합적인 고찰과 명명 규약에 의거한 합법적 절차에 따라 본 분류군의 정당공표를 시도할 것이다(Song and Hong, in prep.).
2. Kirilowii group
Maximowicz (1879)에 의해 처음 S. kirilowii가 인식된 이후, 잎 하면에 존재하는 성상모의 특징으로 S. assurgens와 S. arborea가 구별되었고, Rehder (1913)는 화축과 잎 하면 성상모의 빈도로 S. arborea var. glabrata, S. arborea var. subtomentosa를 기재 및 제시하였다. 하지만, Rahn (1989)은 북위 35°를 경계로 위쪽에 분포하는 S. kirilowii에서는 성상모가 나타나지 않지만, 아래쪽에 분포하는 분류군에서는 드물게 성상모가 산재하는 것을 확인하며, S. kirilowii 한 종으로 통합하여 인식하였다. 한편, Gu and Alexander (2003)는 화서가 느슨하고, 가지가 퍼지며, 과병이 늘어지는 특징뿐만 아니라 서식지 고도의 차이로 S. arborea와 S. kirilowii를 구분하였다. 본 연구 결과, S. arborea와 S. kirilowii 모두에서 화서가 느슨하고, 가지가 펴지는 특징으로 이들 분류군의 식별형질로서 바람직하지 않은 것으로 나타났으며, PCA 결과, 정소엽과 측소엽의 형질과 잎 하면 선점과 성상모의 빈도 등의 영양형질이 식별형질로 유용한 것으로 나타났다.
쉬땅나무속 분류군의 특성 및 분류
Rahn (1989)이 제시한 쉬땅나무속( Sorbaria) 4분류군과 기존 문헌상에서 구분하여 제시한 종 및 종하 분류군을 포함하여, 정량적 형질을 측정하고, 비교 검토하였다.
1. S. grandiflora
본 종은 러시아에 분포하는 분류군으로, 대체로 생식형질이 속 내 다른 분류군에 비해 큰 특징을 지닌다( Maximowicz, 1879; Rahn, 1989). 본 연구 결과, 영양형질로 구분되는 소엽의 개수, 엽맥의 수는 적게 나타났으며, 소엽의 크기 역시 속 내 다른 분류군에 비해 작게 나타났고, 생식형질은 선행 연구에서와 같이 동일하게 속 내 다른 분류군과 비교하여 상대적으로 크게 나타났다. 소엽의 수는 평균 12개에서 최대 15개로 확인되었으며, 정소엽, 측소엽의 크기는 각각 평균 22.4×8.7 mm, 31.1×10.2 mm (길이 ×너비)이었고, 엽맥의 수는 7개에서 최대 15개로, 전반적인 영양형질이 S. sorbifolia complex에 비해 1/2정도로 나타났다. 영양형질을 이용한 PCA 결과에서 정소엽과 측소엽의 크기가 S. grandiflora ( S. rhoifolia)와 S. sorbifolia complex를 구분하는 주요 형질로 확인되었다. 생식형질에서는 화서의 크기가 다른 분류군에 비해 작게 나타나 확연히 구분되었으며, 정량적 형질의 평균이 각각 악편의 크기 2.6×1.8 mm, 화판의 크기 5.4×4.7 mm이고, 꽃의 직경 11.8 mm, 열매의 크기 7.3×5.4 mm (길이×너비)로 S. sorbifolia complex에 비해 1.5–2배 가까이 큰 것으로 확인되었다. PCA 결과, 화판과 꽃의 크기가 주요 식별형질로서 유용함이 검증되었다.
2. S. sorbifolia
본 분류군은 러시아와 중국의 북동 지역(만주)에 분포하며( Rahn, 1989), 일부 북미와 유럽에 식재되어, 토착화(naturalization) 된 것으로 보고되었다( Rudolf, 1974). Maximowicz (1879)는 잎 하면 성상모의 발달 여부로 쉬땅나무( S. sorbifolia var. stellipila)를 구분하여 기재하였고, 이후 Schneider (1905)는 자방의 털의 유무와 악편의 형태 등의 특징으로 구분하여 종 수준으로 승격하여 S. stellipila로 인식하였으며, 종하 분류군으로 S. stellipila var. incerta C. K. Schneid.를 새로이 기재하였다. 이후 Kitagawa (1939)는 S. sorbifolia f. incerta인 품종으로 격하시켜 인식하였다. 본 연구 결과, 종 하 분류군들 대부분의 형질에서 기본종과 형질의 차이 없이 중복되는 결과가 나타났고, 가장 큰 차이를 보이는 형질로 성상모 및 선점의 빈도 형질이 확인되었다. 국내에 분포하고 있는 쉬땅나무( S. sorbifolia var. stellipila)는 성상모의 빈도가 2 mm 2당 42개에서 최대 104개로 뚜렷하게 구분되었으며, 점쉬땅나무로 인식되고 있는 분류군의 선점의 개수는 2 mm 2당 57개에서 최대 135개로 구분되었고, 소엽의 엽맥 수가 다른 분류군에 비해 상대적으로 많아 변종 수준으로 인식함이 바람직하다고 판단되었다. 한편, 청쉬땅나무( S. sorbifolia f. incerta)는 다른 분류군과 뚜렷한 형질 차이가 나타나지 않았다. 따라서, 본 연구에서는 기본종( S. sorbifolia var. sorbifolia)을 포함하여, 성상모의 높은 빈도로 구분되는 쉬땅나무( S. sorbifolia var. stellipila)와 선모의 빈도로 구분되는 점쉬땅나무로 인식하였다.
3. S. kirilowii (좀쉬땅나무)
중국에 자생하는 분류군으로, 최근에는 한국과 일본에도 분포하는 것으로 보고되었다. 한편, 본 종은 중국 고유종인 S. arborea complex와 화서의 밀집도, 가지의 퍼지는 정도, 과병의 직립여부 등의 정성적 형질에 따라 구분되는 것으로 보고된 바 있다( Gu and Alexander, 2003). 하지만, 본 연구 결과, 이들을 구별하는 정성적 형질은 뚜렷한 구분없이 형질의 중첩이 나타나 식별형질로 바람직하지 않은 것으로 판단되었다. 영양형질의 PCA 결과, 정소엽, 측소엽의 크기와 측소엽 엽맥의 수, 선점, 성상모의 빈도의 형질로 구분됨을 확인하였다. 대부분 좀쉬땅나무는 정소엽 44.2×14.5 mm, 측소엽 49.1×15.8 mm (길이 × 너비)이며, 엽맥의 수 20개로 정소엽 54.2×19.2 mm, 측소엽 62.6×16.8 mm (길이×너비)이고, 엽맥의 수 27개인 S. arborea complex에 비해 작게 나타나 구분되었으며, 특히 잎 하면에 선점이 나타나지 않았다. S. arborea complex에서는 선모의 빈도가 높게 나타나는 S. arborea var. arborea와 성상모의 빈도가 높게 나타나는 S. arborea var. subtomentosa로 구분할 수 있었다. 다만, 많은 형질에서 좀쉬땅나무와 형질의 중첩을 보이는 S. arborea var. grabrata는 좀쉬땅나무의 이명으로 처리하는 것이 마땅할 것으로 판단하였다. 특히, 일부 정량적 형질로 좀쉬땅나무와 S. arborea complex가 구분되나, 대부분의 형질과 지리적 분포가 중첩되어, Rahn (1989)이 제시한 종 개념을 지지하였다. 추후, 이들 분류군 간의 종 수준의 인식이 마땅한지 정성적, 정량적 형질의 중첩과 차이, 지리적 분포 및 서식지 정보 등을 종합적으로 고려하여, 종 또는 종 하 분류군으로의 분류학적 재검토가 필요할 것으로 사료된다.
4. S. tomentosa
히말라야 산맥을 중심으로 인도, 파키스탄, 아프가니스칸 등지에 분포하는 분류군으로, 잎의 너비, 엽연의 거치, 털의 유무 및 분포 등의 형질을 기반으로 S. tomentosa var. tomentosa, S. tomentosa var. angustifolia 두 분류군으로 인식하고 있다( Rahn, 1989). 본 연구 결과, S. tomentosa var. tomentosa는 평균적으로 정소엽의 크기가 65.9×20.2 mm이고, 측소엽의 크기가 80.9×22.6 mm이며, 성상모의 빈도가 22.5개/2 mm 2로 확인되었고, S. tomentosa var. angustifolia는 정소엽의 크기가 40.6×8.5 mm이며, 측소엽의 크기가 44.2×8.4 mm이고, 성상모의 빈도가 0개/2 mm 2로 나타나 이들 분류군을 정량적 형질로 쉽게 구분할 수 있었다. 또한, 이중거치가 나타나는 기본종은 엽연 거치의 원측면, 근측면 길이에서 각각 2.4 mm, 5.5 mm로 나타났으며, 단순거치가 나타나는 S. tomentosa var. angustifolia에서는 각각 1.1 mm, 2.3 mm로 나타나, 엽연 거치의 길이도 주요 식별형질로 적용됨을 확인하였다. 본 연구에서는 정량적 형질로 두 분류군으로 뚜렷이 구분할 수 있어, 이들을 변종 수준으로 인식한 Rahn (1989)의 체계를 지지하였다. 결론적으로, 쉬땅나무속은 주요 정성적 형질로 크게 두 그룹으로 구분할 수 있었다. 또한 각 그룹 내에서 분류군은 PCA와 단변량 분석을 통해 주요 식별형질들이 탐색되었고, 확인된 주요 정량적 형질을 이용하여 종 및 종하 분류군으로 구분할 수 있었다. 특히, 종하 분류군을 구분하는 데에는 잎, 소엽의 크기, 엽맥의 수, 털의 빈도 등의 형질이 식별형질로서 유용함을 밝혔다. 본 수리분류학적 연구 결과는 본 속이 포함된 쉬땅나무족의 잎의 해부학적( Song and Hong, 2014), 미세형태학적( Song and Hong, 2016a), 화피의 미세형태학적( Song and Hong, 2016b), 화분학적( Song et al., 2016), 엽병의 해부학적( Song and Hong, 2018) 형질과 분자계통학적 연구 결과를 포함한 종합적인 형질의 고찰과 지리적 분포, 명명규약에 따른 논의가 수반되어, 궁극적으로 본 속의 속 내/ 종 간/ 종 내 분류체계가 확립될 것이다(Song and Hong, in prep.).
ACKNOWLEDGMENTS
The authors would like to thank the directors of the herbaria of A, C, GH, K, KH, KHUS, KWNU, NEBC, MO, PE, and SNUA for permitting the examination of specimens through loans. Two anonymous reviewers, whose comments and corrections improved the work, are also acknowledged. This study is a part of the Ph.D. dissertation of the first author (J.-H. Song) and is supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Sciences and Technology (NRF-2012R1A1A2004149) to S.-P. Hong.
Fig 1.
Univariate statistics with the means and 95% confidence intervals values for the discriminating characters for the genus Sorbaria. SG, S. grandiflora and S. rhoifolia; SS cmx (S. sorbifolia, S. sorbifolia var. stellipila, S. sorbifolia var. glandulosa, S. sorbifolia f. incerta); SK cmx (S. arborea, S. arborea var. glabrata, S. arborea var. subtomentosa, S. kirilowii); ST cmx (S. tomentosa, S. tomentosa var. angustifolia). 
Fig 2.
Schematic representation of plants of the Sorbaria sorbifolia complex and related species with an indication of the measurement of the characters. Keys are given in Table 1. 
Fig 3.
Principal components analysis plot of the Sorbaria sorbifolia complex and related species based on vegetative characters (see Table 1). A. PC 1 vs. PC 2. B. PC 2 vs. PC 3. 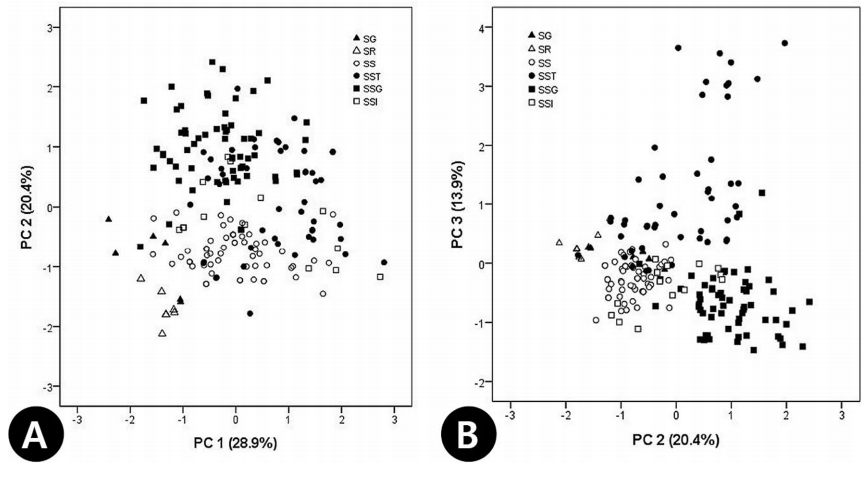
Fig 4.
Principal components analysis plot of the Sorbaria sorbifolia complex and related species based on reproductive characters (see Table 1). 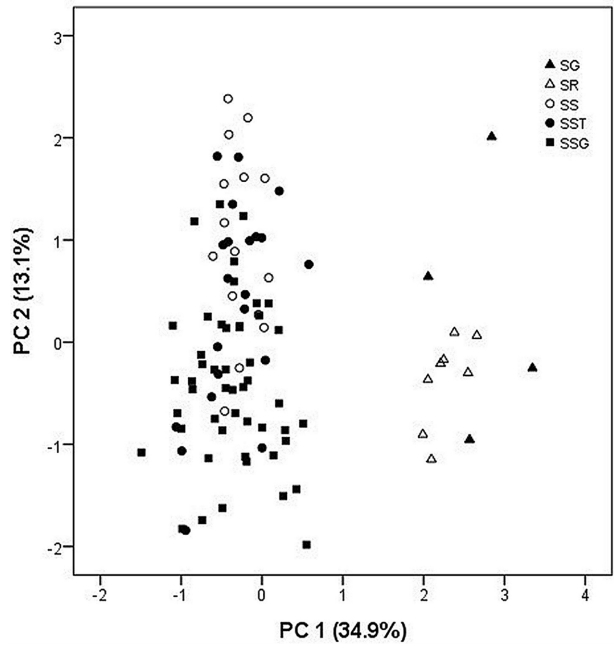
Fig 5.
Univariate statistics with the means and 95% confidence intervals values for the discriminating characters for the Sorbaria sorbifolia complex and related species. SG, S. grandiflora; SR, S. rhoifolia; SSUS, American S. sorbifolia; SSCN, Canadian S. sorbifolia; SSRS, Russian S. sorbifolia; SSCH, Chinese S. sorbifolia; SSTCH, Chinese S. sorbifolia var. stellipila; SSTKR, Korean S. sorbifolia var. stellipila; SSTJP, Japanese S. sorbifolia var. stellipila; SSGKR, Korean S. sorbifolia var. glandulosa; SSIKR, Korean S. sorbifolia f. incerta; SSIJP, Japanese S. sorbifolia f. incerta. 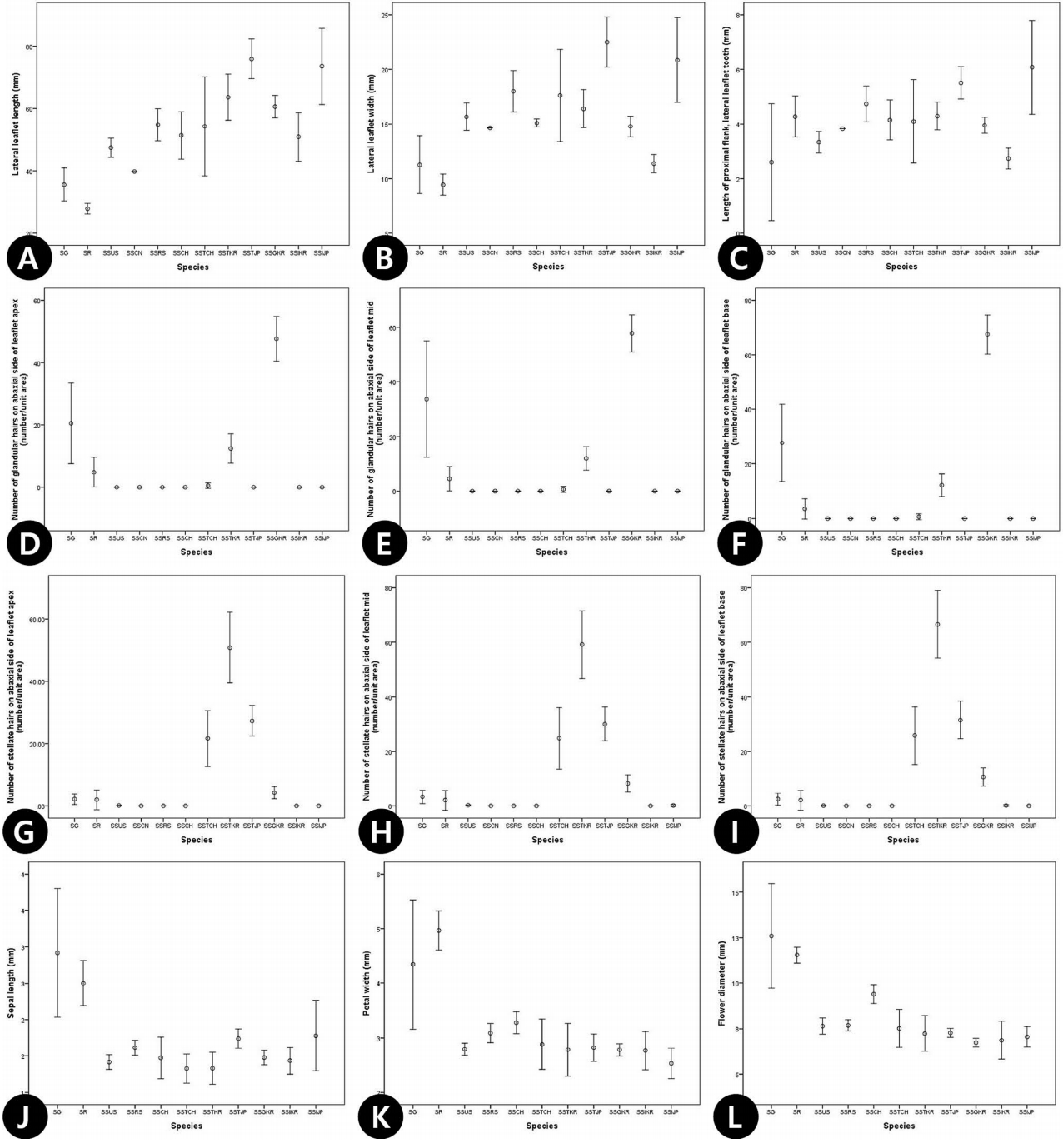
Fig 6.
Schematic representation of plants of the Sorbaria kirilowii complex and related species with an indication of the measurement of the characters. Keys are given in Table 2. 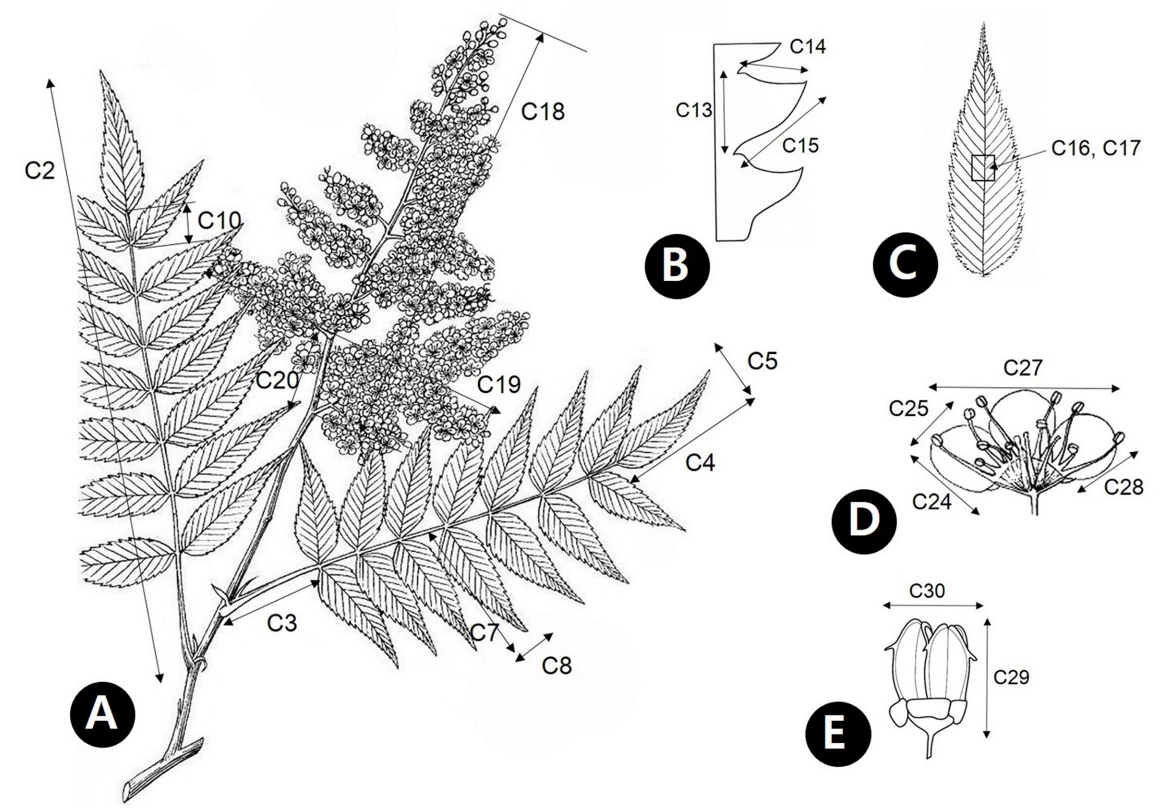
Fig 7.
Principal components analysis (PCA) plot of the Sorbaria kirilowii complex and related species based on vegetative characters (see Table 2). A. PC 1 vs. PC 2. B. PC 2 vs. PC 3. 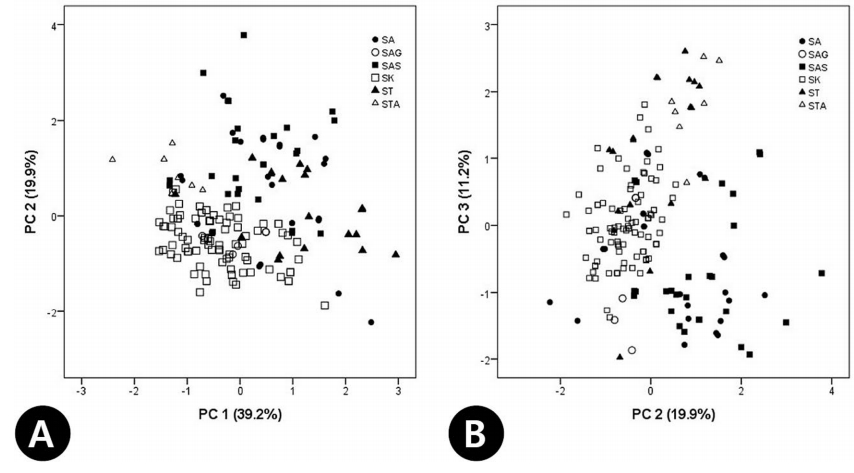
Fig 8.
Principal components analysis (PCA) plot of the Sorbaria kirilowii complex and related species based on reproductive characters (see Table 2). A. PC 1 vs. PC 2. B. PC 2 vs. PC 3. 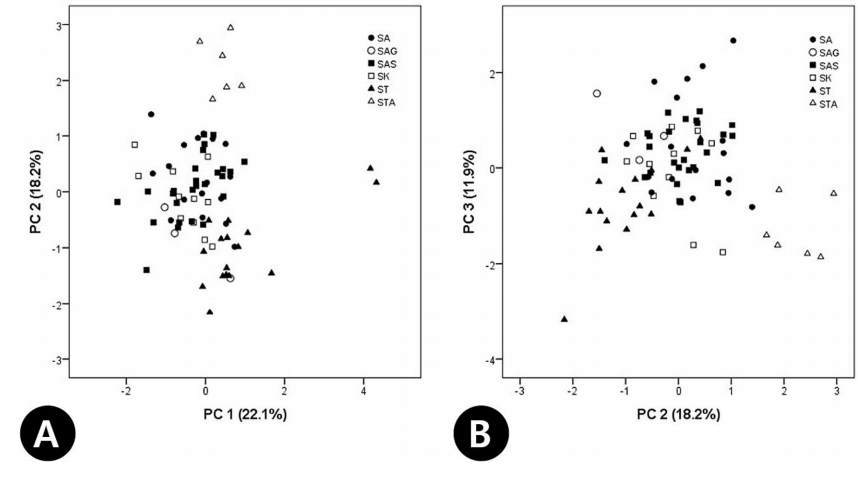
Fig 9.
Univariate statistics with the means and 95% confidence intervals values for the discriminating characters for the Sorbaria kirilowii complex and related species. SA, S. arborea; SAG, S. arborea var. glabrata; SAS, S. arborea var. subtomentosa; SKCH, Chinese S. kirilowii; SKKR, Korean S. kirilowii; ST, S. tomentosa; STA, S. tomentosa var. angustifolia. 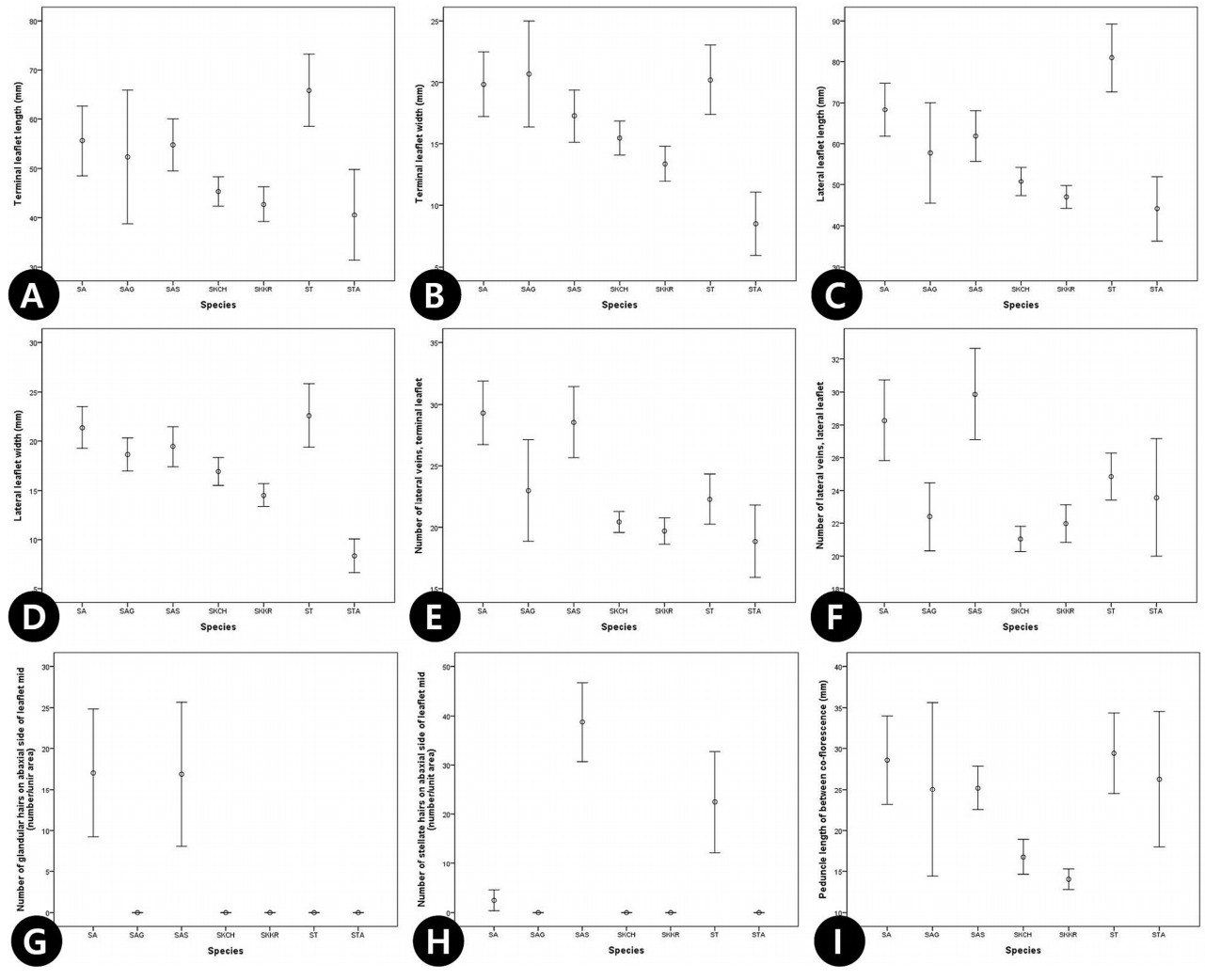
Table 1.
Morphological characters of the Sorbaria sorbifolia complex and related species used in the principal components analysis. Units of measurement are given in parentheses.
|
Vegetative characters |
|
Leaf characters |
|
C1 |
Number of leaflet |
|
C2 |
Length of leaf (mm) |
|
C3 |
Length of petiole (mm) |
|
Leaflet characters |
|
C4 |
Terminal leaflet length (mm) |
|
C5 |
Terminal leaflet width (mm) |
|
C6 |
Ratio of length/width of terminal leaflet |
|
C7 |
Lateral leaflet length (mm) |
|
C8 |
Lateral leaflet width (mm) |
|
C9 |
Ratio of length/width of middle leaflet |
|
C10 |
Petiole length of terminal leaflet (mm) |
|
C11 |
Number of lateral veins, terminal leaflet |
|
C12 |
Number of lateral veins, lateral leaflet |
|
C13 |
Length of tooth, lateral leaflet (mm) |
|
C14 |
Length of distal flank, lateral leaflet tooth (mm) |
|
C15 |
Length of proximal flank, lateral leaflet tooth (mm) |
|
C16 |
Number of glandular hairs on abaxial side of leaflet apex (number/2 mm2) |
|
C17 |
Number of glandular hairs on abaxial side of leaflet mid (number/2 mm2) |
|
C18 |
Number of glandular hairs on abaxial side of leaflet base (number/2 mm2) |
|
C19 |
Number of stellate hairs on abaxial side of leaflet apex (number/2 mm2) |
|
C20 |
Number of stellate hairs on abaxial side of leaflet mid (number/2 mm2) |
|
C21 |
Number of stellate hairs on abaxial side of leaflet base (number/2 mm2) |
|
Reproductive characters |
|
Flower characters |
|
C22 |
Inflorescence length (mm) |
|
C23 |
Inflorescence width (mm) |
|
C24 |
Sepal length (mm) |
|
C25 |
Sepal width (mm) |
|
C26 |
Ratio of length/width of sepal |
|
C27 |
Petal length (mm) |
|
C28 |
Petal width (mm) |
|
C29 |
Ratio of length/width of petal |
|
C30 |
Flower diameter (mm) |
|
C31 |
Filament length (mm) |
|
Fruit characters |
|
C32 |
Fruit length (mm) |
|
C33 |
Fruit width (mm) |
|
C34 |
Ratio of length/width of fruit |
|
Seed characters |
|
C35 |
Seed length (mm) |
|
C36 |
Seed width (mm) |
|
C37 |
Ratio of length/width of seed |
Table 2.
Morphological characters of the Sorbaria kirilowii complex and related species used in the principal components analysis. Units of measurement are given in parentheses.
|
Vegetative characters |
|
Leaf characters |
|
C1 |
Number of leaflet |
|
C2 |
Length of leaf (mm) |
|
C3 |
Length of petiole (mm) |
|
Leaflet characters |
|
C4 |
Terminal leaflet length (mm) |
|
C5 |
Terminal leaflet width (mm) |
|
C6 |
Ratio of length/width of terminal leaflet |
|
C7 |
Lateral leaflet length (mm) |
|
C8 |
Lateral leaflet width (mm) |
|
C9 |
Ratio of length/width of middle leaflet |
|
C10 |
Petiole length of terminal leaflet (mm) |
|
C11 |
Number of lateral veins, terminal leaflet |
|
C12 |
Number of lateral veins, lateral leaflet |
|
C13 |
Length of tooth, lateral leaflet (mm) |
|
C14 |
Length of distal flank, lateral leaflet tooth (mm) |
|
C15 |
Length of proximal flank, lateral leaflet tooth (mm) |
|
C16 |
Number of glandular hairs on abaxial side of leaflet mid (number/2 mm2) |
|
C17 |
Number of stellate hairs on abaxial side of leaflet mid (number/2 mm2) |
|
Reproductive characters |
|
Flower characters |
|
C18 |
Inflorescence length (mm) |
|
C19 |
Inflorescence width (mm) |
|
C20 |
Peduncle length of between co-florescence (mm) |
|
C21 |
Sepal length (mm) |
|
C22 |
Sepal width (mm) |
|
C23 |
Ratio of length/width of sepal |
|
C24 |
Petal length (mm) |
|
C25 |
Petal width (mm) |
|
C26 |
Ratio of length/width of petal |
|
C27 |
Flower diameter (mm) |
|
C28 |
Filament length (mm) |
|
Fruit characters |
|
C29 |
Fruit length (mm) |
|
C30 |
Fruit width (mm) |
|
C31 |
Ratio of length/width of fruit |
|
Seed characters |
|
C32 |
Seed length (mm) |
|
C33 |
Seed width (mm) |
|
C34 |
Ratio of length/width of seed |
Table 3.
Loadings of the first three principal components for 21 vegetative characters of Sorbaria sorbifolia complex and related species. Character codes correspond to those of Table 1. The components that were loaded most highly for each character are in bold.
|
Character codes |
Principal components
|
|
PC1 |
PC2 |
PC3 |
|
C1 |
0.0210 |
0.4670
|
0.1760 |
|
C2 |
0.7740
|
0.3000 |
-0.0600 |
|
C3 |
0.6990
|
0.1130 |
0.1130 |
|
C4 |
0.7520
|
0.0840 |
-0.2500 |
|
C5 |
0.7580
|
-0.2640 |
-0.2190 |
|
C6 |
-0.0960 |
0.5930
|
0.0140 |
|
C7 |
0.8180
|
0.4140 |
-0.0800 |
|
C8 |
0.8660
|
-0.0810 |
-0.0490 |
|
C9 |
-0.0710 |
0.7540
|
-0.0660 |
|
C10 |
0.4960
|
-0.1310 |
-0.2650 |
|
C11 |
0.2600 |
0.5080
|
-0.2120 |
|
C12 |
0.2910 |
0.7210
|
-0.1610 |
|
C13 |
0.7970
|
-0.2660 |
-0.1110 |
|
C14 |
0.6430
|
0.0890 |
-0.1010 |
|
C15 |
0.7680
|
-0.1650 |
-0.0910 |
|
C16 |
-0.2450 |
0.7090
|
-0.3250 |
|
C17 |
-0.2690 |
0.7100
|
-0.3740 |
|
C18 |
-0.2870 |
0.7180
|
-0.3850 |
|
C19 |
0.2630 |
0.3030 |
0.8700
|
|
C20 |
0.2200 |
0.3510 |
0.8530
|
|
C21 |
0.2040 |
0.3790 |
0.8430
|
|
Eigenvalues |
6.0700 |
4.2790 |
2.9330 |
|
Cumulative % of eigenvalues |
28.9 |
49.3 |
63.2 |
Table 4.
Loadings of the first three principal components for 16 reproductive characters of Sorbaria sorbifolia complex and related species. Character codes correspond to those of Table 1. The components that were loaded most highly for each character are in bold.
|
Character codes |
Principal components
|
|
PC1 |
PC2 |
PC3 |
|
C22 |
-0.5810
|
0.4540 |
0.2490 |
|
C23 |
-0.3880 |
0.4820
|
0.4520 |
|
C24 |
0.8470
|
0.2320 |
0.1480 |
|
C25 |
0.6800
|
-0.0280 |
0.1540 |
|
C26 |
0.4670
|
0.4610 |
0.0290 |
|
C27 |
0.8770
|
0.3020 |
0.1540 |
|
C28 |
0.8480
|
0.1590 |
0.1890 |
|
C29 |
0.2780 |
0.4630
|
-0.0690 |
|
C30 |
0.8710 |
0.2270
|
0.0790 |
|
C31 |
-0.0820 |
0.1630 |
0.3570
|
|
C32 |
0.6110
|
-0.3040 |
0.0160 |
|
C33 |
0.6260
|
-0.5260 |
0.4080 |
|
C34 |
-0.1960 |
0.5270 |
-0.5740
|
|
C35 |
-0.4470 |
-0.3430 |
0.4830
|
|
C36 |
0.2870 |
-0.4470
|
-0.2800 |
|
C37 |
-0.5370 |
0.1150 |
0.5960
|
|
Eigenvalues |
5.5910 |
2.0920 |
1.6730 |
|
Cumulative % of eigenvalues |
34.9 |
48.0 |
58.5 |
Table 5.
Loadings of the first three principal components for 17 vegetative characters of Sorbaria kirilowii complex and related species. Character codes correspond to those of Table 2. The components that were loaded most highly for each character are in bold.
|
Character codes |
Principal components
|
|
PC1 |
PC2 |
PC3 |
|
C1 |
-0.3160 |
0.0980 |
0.6400
|
|
C2 |
0.8330
|
0.2560 |
0.3000 |
|
C3 |
0.7270
|
0.1920 |
0.1440 |
|
C4 |
0.8410
|
0.2820 |
0.0990 |
|
C5 |
0.8720
|
-0.1530 |
-0.2710 |
|
C6 |
-0.3090 |
0.6000
|
0.5620 |
|
C7 |
0.8900
|
0.2540 |
0.2270 |
|
C8 |
0.9250
|
-0.0730 |
-0.1880 |
|
C9 |
-0.2270 |
0.4950 |
0.6750
|
|
C10 |
0.7250
|
-0.1170 |
-0.0170 |
|
C11 |
0.2880 |
0.7620
|
-0.3130 |
|
C12 |
0.2700 |
0.8320
|
-0.1960 |
|
C13 |
0.8300
|
-0.2870 |
0.2090 |
|
C14 |
0.3470 |
-0.5970
|
0.3030 |
|
C15 |
0.7030
|
-0.5020 |
0.2990 |
|
C16 |
0.0440 |
0.5880
|
-0.2730 |
|
C17 |
0.3150 |
0.4050
|
-0.0690 |
|
Eigenvalues |
6.6700 |
3.3790 |
1.9030 |
|
Cumulative % of eigenvalues |
39.2 |
59.1 |
70.3 |
Table 6.
Loadings of the first three principal components for 17 reproductive characters of Sorbaria kirilowii complex and related species. Character codes correspond to those of Table 2. The components that were loaded most highly for each character are in bold.
|
Character codes |
Principal components
|
|
PC1 |
PC2 |
PC3 |
|
C18 |
-0.1640 |
-0.3620
|
0.3170 |
|
C19 |
-0.3260 |
-0.3080 |
0.4720
|
|
C20 |
0.1330 |
-0.0930 |
0.4730
|
|
C21 |
0.6570
|
0.2770 |
-0.3260 |
|
C22 |
0.2800 |
-0.3100
|
-0.0750 |
|
C23 |
0.4510 |
0.4630
|
-0.2600 |
|
C24 |
0.8110
|
0.1660 |
0.4560 |
|
C25 |
0.6290
|
0.1490 |
0.5230 |
|
C26 |
0.5950
|
0.1150 |
0.1420 |
|
C27 |
0.6220
|
0.1810 |
0.3630 |
|
C28 |
0.0930 |
0.4390
|
0.4380 |
|
C29 |
-0.0410 |
0.7370
|
0.0120 |
|
C30 |
-0.3120 |
0.8710
|
0.0200 |
|
C31 |
0.5000 |
-0.5020
|
-0.1220 |
|
C32 |
0.3160 |
0.3810 |
-0.5050
|
|
C33 |
-0.4520 |
0.6380
|
0.0690 |
|
C34 |
0.6570
|
-0.2750 |
-0.4500 |
|
Eigenvalues |
3.7510 |
3.0920 |
2.0300 |
|
Cumulative % of eigenvalues |
22.1 |
40.3 |
52.2 |
Literature Cited
Bailey, H. K.. 1949. Sorbaria (Ser.) A. Br. Manual of Cultivated Plants. Most Commonly Grown in the Continental United States and Canada. Revised ed. The Macmillan Company, New York. 501-502.
Ball, P. W.. 1968. Sorbaria (Ser.) A. Br. Flora of Europaea. Vol. 2. Tutin, T. G.. Heywood, V. H.. Burges, N. A.. Valentine, D. H.. Walters, S. M.. Webb, D. A. (eds.), Cambridge University Press, Cambridge. 4 pp.
Braun, A.. 1860. Sorbaria sorbifolia (L.) A. Braun in Ascherson. Flora der Provinz Brandenburg, der Altmark und des Herzogthums Magdeburg. Vol. 1. Ascherson, P. (ed.), Hirschwald, Berlin. 177 pp.
Brouillet, L.. 2015. Sorbarieae. Flora of North America. Vol. 9. Flora of North America Editorial Committee (ed.), Oxford University Press, New York. 392-397.
Chang, C.S. and Kim, H.. 2002. Overlooked and invalidly published names of Korean woody plants. Korean Journal of Plant Taxonomy 32: 363-371 (in Korean).   Ellis, B.. Daly, D.C.. Hickey, L.J.. Mitchell, J.V.. Johnson, K.R.. Wilf, P. and Wing, S.L.. 2009. Manual of Leaf Architecture. Cornell University Press, Ithaca, NY. 216 pp.
Gu, C. and Alexander, C.. 2003. Sorbaria. Flora of China. Vol. 9. Wu, Z. Y.. Raven, P. H.. Hong, D. Y. (eds.), Science Press, Beijing and Missouri Botanical Garden Press, St. Louis, MO. 75-76.
Henrickson, J.. 2015. Sorbaria (Seringe) A. Braun in P. F. A. Ascherson. Flora of North America. Vol. 9. Flora of North America Editorial Committee (ed.), Oxford University Press, New York. 396-397.
Ikeda, H.. 2001. Sorbaria (Ser.) A. Br. ex Aschers. Flora of Japan. Vol. IIb. Angiospermae; Dicotyledoneae; Archichlamideae (b). Iwatsuki, K.. Yamazaki, T.. Boufford, D. E.. Ohba, H. (eds.), Kodansha, Ltd., Tokyo. 102 pp.
Judd, W.S.. Campbell, C.S.. Kellogg, E.A.. Stevens, P.F. and Donoghue, M.J.. 1999. Plant Systematics: a Phylogenetic Approach. Sinauer Associates, Sunderland, MA. 576 pp.
Kaburaki, N.. 1940. Chosen Sinrin Zumoku Yoran, Vol. 1. Mansen Zituyo Ringyo Binran, 141. Yogendo, Tokyo. 123 (in Japanese).
Kim, H.. Lee, H.S.. Park, S. and Chang, C.S.. 2005. Invalid names published by T. B. Lee. Korean Journal of Plant Taxonomy 35: 211-226 (in Korean). Kitagawa, M.. 1939. Sorbaria sorbifolia f. incerta (Schneider) Kitagawa. Lineam. Fl. Mansh. 275.
Lee, S.T.. 2007. Spiraeoideae, Sorbaria. The Genera of Vascular Plants of Korea. Flora of Korean Editorial Committee (ed.), Academy Publishing Co., Seoul. 533, 540–541.
Lee, T.B.. 1980. Illustrated Flora of Korea. Hyangmoon Co., Seoul. 426 (in Korean).
Lingdi, L.. Cuizhi, G.. Chaoluan, L.. Alexander, C.. Bartholomew, B.. Brach, A.R.. Boufford, D.E.. Ikeda, H.. Ohba, H.. Robertson, K.R. and Spongberg, S.A.. 2003. Rosaceae. Flora of China, Vol. 9. Wu, Z. Y.. Raven, P. H.. Hong, D. Y. (eds.), Science Press, Beijing and Missouri Botanical Garden Press, St. Louis, MO. 46-47.
Linnaeus, C.. 1753. Species Plantarum I. Stockholm. 490 pp.
Mabberley, D.J.. 1990. The Plant Book: a Portable Dictionary of the Higher Plants. Cambridge University Press, Cambridge. 546 pp.
Maximowicz, C.J.. 1879. Sorbaria. Acta Horti Petropolitani 6: 222-225.
McNeill, J.. Barrie, F.R.. Buck, W.R.. Demoulin, V.. Greuter, W.. Hawksworth, D.L.. Herendeen, P.S.. Knapp, S.. Marhold, K.. Prado, J.. Prud'homme Van Reine, W.. Smith, G.F.. Wiersema, J.H. and Turland, N.J.. 2012. International Code of Nomenclature for Algae, Fungi, and Plants (Melbourne Code) adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011. Koeltz Scientific Books, Konigstein. 208 pp.
Pojarkova, A.I.. 1971. Sorbaria. A. Br. Flora of the U.S.S.R., Vol. IX. Komarov, V. L.. Yuzepchuk, S. V. (eds.), U.S. Department of Commerce, Springfield, VA. 240-243 (in English edition).
Purohit, K.M. and Panigrahi, G.. 1984. Taxonomic studies on Sorbaria (Rosaceae) in India. Indian Journal of Forestry 7: 207-213.
Rahn, K.. 1989. A survey of the genus Sorbaria (Rosaceae). Nordic Journal of Botany 8: 557-563. Rehder, A.. 1913. Sorbaria A. Br. Plantae Wilsonianae. Vol. 1. Sargent, C. S. (ed.), The University Press, Cambridge. 47-48.
Rehder, A.. 1940. Sorbaria A. Br. Manual of Cultivated Trees and Shrubs. Macmillian Company, New York. 343-345.
Rudolf, P. O.. 1974. Sorbaria sorbifolia (L.) A. Br. Ural False Spiraea. Agriculture Handbook No. 450. U. S. Department of Agriculture, Washington, DC. 779 pp.
Schneider, C.K.. 1905. Illustriertes Handbuch der Laubholzkunde. Vol. 1. Verlag von Gustav Fisher, Jena. 486-491.
Schönbeck-Temesy, E.. 1969. Sorbaria (Ser.) A. Br. Flora Iranica. Vol. 66. Rechinger, K. H. (ed.), Akademische Druckund Verlagsanstalt, Graz. 10-12.
Seringe, N.C.. 1825. Rosaceae. Prodromus Systematis Naturalis Regni Vegetabilis. De Candolle, A. P. (ed.), Treuttel et Würtz, Paris. 525-639.
Song, J.-H. and Hong, S.-P.. 2014. The taxonomic implication of leaf anatomy in tribe Sorbarieae (Spiraeoideae: Rosaceae). Korean Journal of Plant Taxonomy 44: 119-131 (in Korean). Song, J.-H. and Hong, S.-P.. 2016a. Taxonomic significance of the leaf micromorphology in the tribe Sorbarieae (Spiraeoideae: Rosaceae). Korean Journal of Plant Taxonomy 46: 199-212 (in Korean). Song, J.-H. and Hong, S.-P.. 2016b. A study on the petal micromorphological characteristics of the tribe Sorbarieae (Rosaceae). Korean Journal of Plant Resources 29: 376-384 (in Korean). Song, J.-H. and Hong, S.-P.. 2018. Comparative petiole anatomy of the tribe Sorbarieae (Rosaceae) provide new taxonomically informative characters. Nordic Journal of Botany 36: e01702. https://doi.org/10.1111/njb.01702. Song, J.-H.. Chang, C.-S. and Hong, S.-P.. 2010. First record of Sorbaria kirilowii (Rosaceae-Spiraeoideae) from Korea. Korean Journal of Plant Taxonomy 40: 174-178 (in Korean). Song, J.-H.. Moon, H.-K. and Hong, S.-P.. 2016. Pollen morphology of the tribe Sorbarieae (Rosaceae). Plant Systematics and Evolution 302: 853-869. Thiers, B.. 2018. (continuously updated) Index Herbariorum: a global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium; Retrieved Mar. 10, 2018, available from http://sweetgum.nybg.org/ih/. Tomaszewski, D.. 2001. Sorbaria species cultivated in Poland. Dendrobiology 46: 59-64.
Willis, J.C.. 1948. A Dictionary of the Flowering Plants and Ferns. 6th ed. (Revised). Cambridge University Press, Cambridge. 614 pp.
Yü, T. T. and Ku, T. C.. 1974. Sorbaria (Ser.) A. Br. Flora Reipublicae Popularis Sinicae. Vol. 36. Yü, T. T.. Lu, L. T.. Ku, T. C.. Kuau, K. C.. Chiang, W. F. (eds.), Institutum Botanicum Pekinense Academiae Sinicae, Peking. 75-80 (in Chinese).
APPENDICES
Appendix 1.
Comparison of morphological characters among Sorbaria sorbifolia complex and related species. Character codes correspond to those of Table 1.
|
Character codes |
S. grandiflora
|
S. sorbifolia var. sorbifolia
|
S. sorbifolia var. stellipila
|
S. sorbifolia var. gladulosa
|
S. sorbifolia f. incerta
|
|
C1 |
9.0 (12.1±2.03) 15.0 |
11.0 (16.7±3.8) 29.0 |
11.0 (17.1±3.1) 23.0 |
9.0 (17.2±3.1) 25.0 |
11.0 (18.2±5.0) 23.0 |
|
C2 |
68.5 (109.6±24.4) 140.6 |
141.0 (206.3±37.2) 310.0 |
138.1 (228.7±51.9) 355.0 |
110.0 (202.2±43.1) 290.0 |
165.0 (237.3±49.8) 340.0 |
|
C3 |
14.6 (26.1±7.70) 42.0 |
11.6 (35.2±10.2) 62.8 |
21.5 (42.8±9.2) 68.2 |
15.4 (33.4±8.9) 51.4 |
17.9 (41.6±15.4) 72.3 |
|
C4 |
14.9 (22.4±4.14) 26.8 |
14.0 (47.3±11.9) 72.7 |
28.8 (49.9±14.0) 83.8 |
22.5 (45.2±9.9) 68.0 |
39.8 (56.6±18.7) 96.7 |
|
C5 |
5.9 (8.7±2.1) 12.4 |
9.1 (16.5±4.6) 30.7 |
7.8 (16.3±5.0) 25.4 |
5.0 (13.1±3.4) 22.2 |
9.8 (16.4±6.7) 27.4 |
|
C6 |
1.8 (2.7±0.6) 3.7 |
0.8 (2.9±0.55) 4.0 |
1.9 (3.2±0.7) 4.8 |
2.4 (3.5±0.6) 4.7 |
2.6 (3.6±0.6) 4.9 |
|
C7 |
25.3 (31.1±5.3) 41.7 |
29.5 (50.2±9.7) 80.4 |
32.8 (67.8±16.7) 92.2 |
25.2 (60.6±14.1) 104.0 |
39.2 (61.4±15.9) 87.5 |
|
C8 |
6.9 (10.2±2.0) 13.1 |
10.7 (16.4±3.5) 26.1 |
9.6 (19.2±5.3) 31.2 |
8.6 (14.2±3.8) 25.1 |
9.9 (15.8±5.6) 27.4 |
|
C9 |
2.3 (3.1±0.4) 4.2 |
1.7 (3.0±0.3) 3.8 |
2.5 (3.6±0.6) 4.6 |
2.8 (4.2±0.7) 6.1 |
2.8 (4.1±0.8) 5.3 |
|
C10 |
1.8 (6.1±2.2) 10.5 |
2.1 (8.5±3.0) 15.9 |
0.9 (7.9±3.4) 17.8 |
2.6 (7.3±2.7) 12.3 |
2.9 (7.9±2.9) 13.3 |
|
C11 |
7.0 (9.9±2.1) 13.0 |
13.0 (19.7±3.1) 28.0 |
9.0 (19.3±5.5) 36.0 |
12.0 (21.6±4.2) 30.0 |
15.0 (21.7±3.1) 27.0 |
|
C12 |
10.0 (12.6±1.7) 15.0 |
18.0 (21.7±2.8) 31.0 |
12.0 (23.8±6.3) 39.0 |
15.0 (26.6±3.9) 35.0 |
18.0 (22.9±2.8) 27.0 |
|
C13 |
1.98 (2.8±0.5) 3.7 |
1.0 (3.1±1.0) 5.5 |
2.0 (3.6±1.1) 7.3 |
1.4 (2.8±0.8) 4.8 |
2.2 (3.6±1.5) 6.7 |
|
C14 |
0.87 (1.7±0.7) 2.9 |
0.7 (1.8 ±0.6) 3.2 |
1.3 (2.2±0.6) 4.2 |
1.1 (2.2±0.7) 4.3 |
0.8 (2.0±1.0) 4.4 |
|
C15 |
0.67 (3.6±1.6) 5.2 |
1.1 (3.9±1.3) 6.8 |
2.9 (4.8±1.4) 8.8 |
1.2 (3.9±1.2) 8.1 |
2.2 (4.3±2.1) 9.2 |
|
C16 |
0.0 (11.5±11.8) 35.0 |
0.0 (0.0±0.0) 0.0 |
0.0 (5.5±9.2) 34.0 |
10.0 (47.6±27.9) 126.0 |
0.0 (0.0±0.0) 0.0 |
|
C17 |
0.0 (17.0±19.9) 60.0 |
0.0 (0.0±0.0) 0.0 |
0.0 (5.3±8.5) 35.0 |
14.0 (57.8±26.7) 128.0 |
0.0 (0.0±0.0) 0.0 |
|
C18 |
0.0 (13.9±15.3) 42.0 |
0.0 (0.0±0.0) 0.0 |
0.0 (5.4±8.4) 25.0 |
24.0 (67.4±28.1) 135.0 |
0.0 (0.0±0.0) 0.0 |
|
C19 |
0.0 (2.1±2.9) 10.0 |
0.0 (0.1±0.2) 1.0 |
8.0 (36.9±21.9) 93.0 |
0.0 (4.3±7.8) 42.0 |
0.0 (0.0±0.0) 0.0 |
|
C20 |
0.0 (2.6±3.5) 12.0 |
0.0 (0.1±0.3) 1.0 |
8.0 (42.2±25.3) 102.0 |
0.0 (8.3±12.2) 51.0 |
0.0 (0.1±2.5) 1.0 |
|
C21 |
0.0 (2.3±3.4) 12.0 |
0.0 (0.0±0.3) 1.0 |
10.0 (46.2±27.6) 104.0 |
0.0 (10.7±13.1) 56.0 |
0.0 (0.1±2.5) 1.0 |
|
C22 |
17.7 (44.0±12.6) 63.6 |
80.9 (198.2±61.2) 305.1 |
76.9 (166.4±70.0) 320.0 |
83.3 (163.4±52.3) 290.0 |
130.0 (195.5±42.5) 260.0 |
|
C23 |
22.7 (36.9±7.9) 45.5 |
22.7 (68.4±20.1) 112.8 |
30.3 (61.3±22.4) 110.0 |
21.2 (64.8±21.2) 140.0 |
44.6 (66.9±14.2) 88.1 |
|
C24 |
1.9 (2.6±0.4) 3.4 |
0.9 (1.5±0.3) 2.2 |
0.8 (1.5±0.4) 2.4 |
0.7 (1.5±0.3) 2.2 |
1.0 (1.6±0.4) 2.5 |
|
C25 |
0.9 (1.8±0.4) 2.4 |
0.7 (1.3±0.2) 1.9 |
0.8 (1.2±0.2) 1.9 |
0.8 (1.3±0.2) 1.9 |
1.0 (1.4±0.3) 1.9 |
|
C26 |
1.1 (1.5±0.3) 2.3 |
0.8 (1.2±0.2) 1.8 |
0.8 (1.3±0.3) 2.2 |
0.8 (1.1±0.2) 1.4 |
1.0 (1.1±0.2) 1.5 |
|
C27 |
4.0 (5.4±0.6) 6.5 |
2.5 (3.3±0.6) 5.9 |
1.3 (3.0±0.6) 4.3 |
2.0 (2.9±0.5) 3.7 |
2.3 (2.8±0.3) 3.4 |
|
C28 |
3.4 (4.7±0.6) 5.5 |
2.2 (2.9±0.4) 3.8 |
1.4 (2.8±0.6) 3.7 |
2.0 (2.8±0.4) 3.7 |
2.2 (2.7±0.4) 3.2 |
|
C29 |
0.9 (1.2±0.2) 1.8 |
0.0 (1.1±0.2) 2.1 |
0.8 (1.1±0.1) 1.5 |
1.0 (1.1±0.1) 1.3 |
0.9 (1.1±0.1) 1.3 |
|
C30 |
10.2 (11.8±1.1) 14.3 |
5.9 (7.8±1.0) 10.1 |
5.2 (7.3±1.0) 9.6 |
5.1 (6.7±0.9) 8.7 |
5.3 (6.9±1.0) 8.2 |
|
C31 |
4.3 (5.5±1.3) 8.8 |
2.9 (5.8±1.1) 9.0 |
3.6 (5.5±1.2) 8.4 |
4.1 (6.4±0.9) 8.3 |
3.6 (5.4±1.0) 7.1 |
|
C32 |
5.8 (7.3±0.8) 8.5 |
4.3 (5.1±0.5) 6.1 |
4.6 (5.7±0.5) 6.8 |
4.1 (5.6±0.8) 7.7 |
- |
|
C33 |
3.9 (5.4±0.8) 7.1 |
2.0 (2.9±0.6) 4.0 |
2.3 (3.3±0.5) 4.3 |
2.6 (3.9±0.6) 5.3 |
- |
|
C34 |
0.9 (1.3±0.3) 2.1 |
1.5 (1.8±0.3) 2.2 |
1.3 (1.8±0.3) 2.4 |
1.2 (1.5±0.2) 1.9 |
- |
|
C35 |
2.7 (3.6±0.6) 4.5 |
3.0 (4.6±0.8) 5.7 |
3.2 (4.3±0.5) 5.3 |
3.8 (4.8±0.5) 6.1 |
- |
|
C36 |
0.4 (0.6±0.1) 0.8 |
0.4 (0.5±0.1) 0.7 |
0.3 (0.5±0.1) 0.6 |
0.3 (0.6±0.1) 0.8 |
- |
|
C37 |
5.1 (6.6±0.1) 8.7 |
6.3 (8.7±1.4) 10.9 |
6.5 (9.1±1.6) 12.2 |
5.1 (9.1±1.8) 14.8 |
- |
Appendix 2.
Comparison of morphological characters among Sorbaria kirilowii complex and related species. Character codes correspond to those of Table 2.
|
Character codes |
S. arborea var. arborea
|
S. arborea var. glabrata
|
S. arborea var. subtomentosa
|
S. kirilowii
|
S. tomentosa var. tomentosa
|
S. tomentosa var. angustifolia
|
|
C1 |
11.0 (14.5±2.1) 17.0 |
11.0 (13.3±1.5) 15.0 |
11.0 (14.3±2.2) 17.0 |
9.0 (15.1±2.1) 21.0 |
9.0 (15.1±2.4) 19.0 |
13.0 (16.6±2.7) 21.0 |
|
C2 |
120.1 (236.7±46.7) 310.0 |
150.0 (221.7±52.3) 260.0 |
135.0 (226.7±46.9) 300.0 |
110.0 (188.1±38.4) 290.0 |
130.0 (289.3±60.6) 360.0 |
107.9 (191.8±42.1) 230.0 |
|
C3 |
22.0 (47.4±14.7) 67.3 |
22.7 (29.8±10.6) 51.3 |
19.8 (39.7±11.1) 54.4 |
18.6 (33.4±8.5) 53.0 |
15.4 (56.5±19.6) 93.5 |
17.9 (32.1±7.6) 39.2 |
|
C4 |
25.3 (55.6±17.6) 96.5 |
33.2 (52.3±12.9) 66.8 |
26.9 (54.8±13.8) 84.3 |
23.2 (44.2±10.7) 71.5 |
28.6 (65.9±15.7) 88.2 |
25.8 (40.6±9.9) 57.1 |
|
C5 |
7.6 (19.8±6.5) 36.2 |
15.3 (20.7±4.1) 27.6 |
10.2 (17.3±5.6) 30.6 |
7.1 (14.5±4.7) 28.1 |
7.7 (20.2±6.0) 32.9 |
4.9 (8.5±2.8) 12.2 |
|
C6 |
1.7 (2.9±0.8) 4.0 |
1.9 (2.6±0.6) 3.4 |
2.3 (3.3±0.9) 5.5 |
2.1 (3.2±0.5) 4.9 |
2.3 (3.4±0.7) 5.0 |
4.0 (4.9±0.6) 5.9 |
|
C7 |
40.4 (68.3±16.1) 94.4 |
38.9 (57.8±11.7) 69.5 |
34.9 (61.9±16.1) 87.8 |
33.1 (49.1±10.7) 77.9 |
50.5 (80.9±17.7) 110.5 |
29.7 (44.2±8.5) 58.1 |
|
C8 |
11.9 (21.4±5.3) 37.7 |
15.7 (18.6±1.6) 19.6 |
11.9 (19.5±5.4) 29.6 |
9.1 (15.8±4.5) 29.8 |
11.8 (22.6±6.9) 40.2 |
4.7 (8.4±1.8) 10.5 |
|
C9 |
2.1 (3.2±0.5) 3.9 |
2.5 (3.1±0.4) 3.5 |
2.6 (3.2±0.6) 4.7 |
2.2 (3.2±0.5) 4.5 |
2.2 (3.8±0.9) 5.3 |
4.4 (5.4±0.7) 6.3 |
|
C10 |
5.8 (12.8±5.1) 27.1 |
2.2 (5.8±2.2) 8.5 |
1.2 (10.7±5.4) 21.5 |
3.1 (8.6±3.0) 15.9 |
3.2 (11.8±6.6) 23.4 |
3.2 (6.7±2.4) 9.9 |
|
C11 |
20.0 (29.3±6.4) 43.0 |
19.0 (23.0±3.3) 26.0 |
17.0 (28.6±7.6) 46.0 |
11.0 (20.1±3.1) 27.0 |
18.0 (22.3±4.4) 32.0 |
15.0 (18.9±3.1) 23.0 |
|
C12 |
18.0 (28.3±6.1) 37.0 |
20.0 (22.4±1.7) 24.0 |
17.0 (29.9±7.0) 40.0 |
14.0 (21.5±3.1) 30.0 |
21.0 (24.9±3.0) 31.0 |
18.0 (23.6±3.9) 29.0 |
|
C13 |
0.9 (3.6±1.7) 7.3 |
2.5 (3.3±0.7) 3.9 |
1.5 (2.7±0.8) 4.4 |
1.3 (2.7±0.9) 5.9 |
2.3 (4.6±1.1) 6.4 |
1.1 (2.1±0.4) 2.5 |
|
C14 |
0.9 (1.7±0.6) 3.6 |
1.0 (1.9±1.0) 3.0 |
0.6 (1.3±0.5) 2.5 |
0.8 (2.1±0.6) 3.6 |
0.9 (2.4±1.0) 4.2 |
0.7 (1.1±0.3) 1.6 |
|
C15 |
1.1 (4.2±2.1) 7.5 |
2.3 (4.1±1.3) 5.3 |
1.6 (3.3±1.0) 5.6 |
1.9 (4.2±1.2) 8.6 |
2.3 (5.5±1.7) 9.0 |
1.3 (2.3±0.6) 3.3 |
|
C16 |
0.0 (17.0±19.3) 90.0 |
0.0 (0.0±0.0) 0.0 |
0.0 (16.9±22.2) 90.0 |
0.0 (0.0±0.0) 0.0 |
0.0 (0.0±0.0) 0.0 |
0.0 (0.0±0.0) 0.0 |
|
C17 |
0.0 (2.5±5.1) 22.0 |
0.0 (0.0±0.0) 0.0 |
8.0 (38.7±20.2) 83.0 |
0.0 (0.0±0.0) 0.0 |
2.0 (22.5±21.9) 82.0 |
0.0 (0.0±0.0) 0.0 |
|
C18 |
97.9 (226.1±71.3) 400.0 |
160.0 (210±48.5) 270.0 |
138.4 (246.9±76.2) 360.0 |
100.0 (170.6±36.6) 240.0 |
136.6 (252.5±71.1) 370 |
130.0 (198.5±61.3) 250.0 |
|
C19 |
104.8 (197.6±75.0) 350.0 |
100.0 (128.0±32.7) 180.0 |
54.2 (169.3±66.6) 290.0 |
57.5 (151.0±45.9) 270.0 |
64.2 (118.3±37.5) 220.0 |
33.1 (67.8±35.8) 104.8 |
|
C20 |
13.8 (28.6±13.1) 52.3 |
15.7 (25.0±8.5) 31.3 |
14.4 (25.2±6.6) 40.5 |
6.1 (15.5±6.1) 34.3 |
14.6 (29.4±9.9) 46.1 |
17.5 (26.3±7.9) 33.2 |
|
C21 |
0.5 (0.8±0.2) 1.2 |
0.5 (0.9±0.4) 1.6 |
0.4 (0.7±0.1) 1.0 |
0.3 (0.8±0.2) 1.3 |
0.6 (0.9±0.2) 1.5 |
1.0 (1.1±0.1) 1.3 |
|
C22 |
0.7 (1.1±0.2) 1.5 |
1.0 (1.3±0.3) 1.8 |
0.7 (1.1±0.2) 1.6 |
0.7 (1.1±0.2) 1.9 |
0.9 (1.2±0.2) 1.7 |
0.9 (1.1±0.1) 1.2 |
|
C23 |
0.4 (0.7±0.1) 1.0 |
0.5 (0.7±0.1) 0.9 |
0.4 (0.7±0.1) 1.0 |
0.3 (0.7±0.1) 1.0 |
0.4 (0.8±0.2) 1.1 |
0.9 (1.0±0.1) 1.2 |
|
C24 |
1.6 (2.3±0.3) 2.9 |
2.1 (2.7±0.5) 3.3 |
1.9 (2.4±0.3) 3.0 |
1.3 (2.7±0.6) 5.7 |
1.5 (2.5±0.7) 4.2 |
2.2 (2.5±0.2) 2.8 |
|
C25 |
1.6 (2.3±0.3) 2.9 |
2.1 (2.6±0.5) 3.3 |
1.8 (2.4±0.3) 3.0 |
1.5 (2.7±0.4) 4.1 |
1.7 (2.4±0.4) 3.3 |
2.1 (2.4±0.2) 2.6 |
|
C26 |
0.7 (0.9±0.1) 1.2 |
1.0 (1.0±0.1) 1.1 |
0.9 (1.0±0.1) 1.2 |
0.7 (1.0±0.2) 2.1 |
0.8 (1.0±0.1) 1.3 |
1.0 (1.0±0.1) 1.1 |
|
C27 |
0.9 (5.8±1.3) 7.1 |
6.0 (7.3±1.6) 9.8 |
5.1 (6.3±0.8) 7.6 |
4.3 (6.5±0.8) 8.7 |
4.8 (6.2±1.1) 9.4 |
6.0 (6.5±0.3) 6.9 |
|
C28 |
1.8 (2.9±0.7) 4.8 |
2.1 (2.4±0.4) 2.8 |
1.6 (2.6±0.6) 4.1 |
1.3 (2.7±0.7) 5.0 |
1.3 (2.0±0.5) 3.2 |
2.1 (2.4±0.2) 2.7 |
|
C29 |
2.7 (3.8±0.6) 4.9 |
2.2 (2.9±0.6) 3.7 |
2.6 (3.7±0.4) 4.6 |
3.1 (3.7±0.4) 4.5 |
2.4 (3.2±0.5) 4.2 |
4.5 (4.9±0.2) 5.3 |
|
C30 |
1.2 (2.5±0.6) 3.7 |
1.5 (1.8±0.3) 2.3 |
2.1 (2.6±0.3) 3.2 |
2.2 (2.6±0.3) 3.2 |
1.5 (1.8±2.0) 2.4 |
3.4 (3.8±0.3) 4.1 |
|
C31 |
1.2 (1.5±0.3) 2.6 |
1.3 (1.6±0.3) 2.2 |
1.1 (1.4±0.2) 1.8 |
1.2 (1.4±0.1) 1.6 |
1.5 (1.7±0.2) 2.2 |
1.1 (1.3±0.1) 1.4 |
|
C32 |
2.1 (2.8±0.4) 3.4 |
2.0 (2.5±0.4) 3.1 |
2.0 (2.9±0.4) 1.8 |
2.4 (3.0±0.3) 3.3 |
2.5 (3.3±0.3) 3.7 |
3.8 (4.1±0.2) 4.3 |
|
C33 |
0.3 (1.4±0.1) 0.6 |
0.3 (0.4±0.1) 0.5 |
0.3 (0.5±0.1) 0.6 |
0.3 (0.5±0.1) 0.6 |
0.2 (0.4±0.1) 0.6 |
0.4 (0.6±0.1) 0.6 |
|
C34 |
4.8 (6.3±1.0) 7.9 |
4.1 (6.1±1.9) 9.2 |
4.5 (6.3±1.1) 9.0 |
4.9 (6.5±1.4) 9.7 |
5.3 (8.5±1.6) 11.5 |
6.2 (7.4±0.8) 8.5 |
|
|











