The genus Asplenium L. (Aspleniaceae) includes about 700 taxa worldwide, and 15−21 taxa of Asplenium are known to be distributed in Korea (Park, 1975; Lee, 1980; Lee, 2006; Kim and Sun, 2007; Lee and Lee, 2015). This genus is well characterized by rhizome erect or shortly creeping, and densely scaly throughout or on stipe base, fronds clustered or remote, lamina simple to 4-pinnate and costa usually with all basal basiscopic veins present, linear indisium, echinate spore, n = 36 (Murakami and Schaal, 1994; Iwatsuki, 1995; Lin and Viane, 2012).
Hybridization in natural biological processes occurs frequently among closely related species, and leads to plant speciation (Rieseberg and Ellstrand, 1993; Rieseberg and Wendel, 1993; Arnold, 1997; Rieseberg, 1997).
Interspsecific gene flow may proceed only to the formation of F1 due to factors such as hybrid sterility. If hybrids reproduce, they may backcross with parents at the site of hybridization and create a hybrid swarm. Long-term hybrid swarms can be maintained in different ways (Anderson, 1949; Arnold, 1997). Natural hybridization between parental taxa occurs at a sufficiently high frequency (Ellstrand et al., 2007). Documentation of hybridization has traditionally been based on morphological characters that are intermediate between parental types or that combine parental characters (Grant, 1981; Rieseberg and Ellstrand, 1993). Natural hybridization occurs frequently in Asplenium (Lovis, 1973; Kramer, 1990; Iwatsuki, 1995). Hybrids can be identified easily by their aborted spores and intermediate morphology (Reichstein, 1981; Lovis and Reichstein, 1985; Jessen, 1995).
Asian walking fern, Asplenium ruprechtii Sa. Kurata, produces a sterile hybrid or fertile allotetraploid hybrid with A. incisum Thunb. (Lovies et al., 1972; Ching and Iwatsuki, 1982). It is known to form a hybrid with A. incisum as A. castaneo-viride Baker from Korea (Kwon et al., 2009). In addition to the above hybrid taxon, we newly found two new natural hybrids and an unrecorded hybrid with A. pekinense Hance, A. trichomanes L., A. incisum, and A. sarelii Hooker among 19 taxa in Korea. One hybrid taxon, Asplenium × uiryeongse, nom. nov., with about 20 individuals per 2 m2, was collected from a forest in Uiryeong-gil, Mt. Bukansan, Seoul. The scientific name was assigned based on its habitat, and the local name was designated as ‘geo-mi-sa-cheol-go-sa-ri’ based on its parent names. The other one, A. × montanus, nom. nov., with 50 individuals per 2 m2 , was collected from a forest of Mt. Seongsan in Yeoncheon-gun, Gyeonggi-do. The scientific name was assigned based on its habitat, and the local name was also designated as ‘san-kko-ri-go-sa-ri’ from based its habitat. One unrecorded taxon, A. × kitazawae Kurata & Hutoh, with about 30 individuals per 2 m2 , was collected from Do-dong, Daegu-si. The local name was designated as ‘geo-mi-dol-dam-go-sa-ri’ based on its parent names.
We compared and analyzed morphological characters between Asplenium hybrid taxa and related taxa of Asplenium in order to elucidate their taxonomic relationship.
Taxonomic Treatment
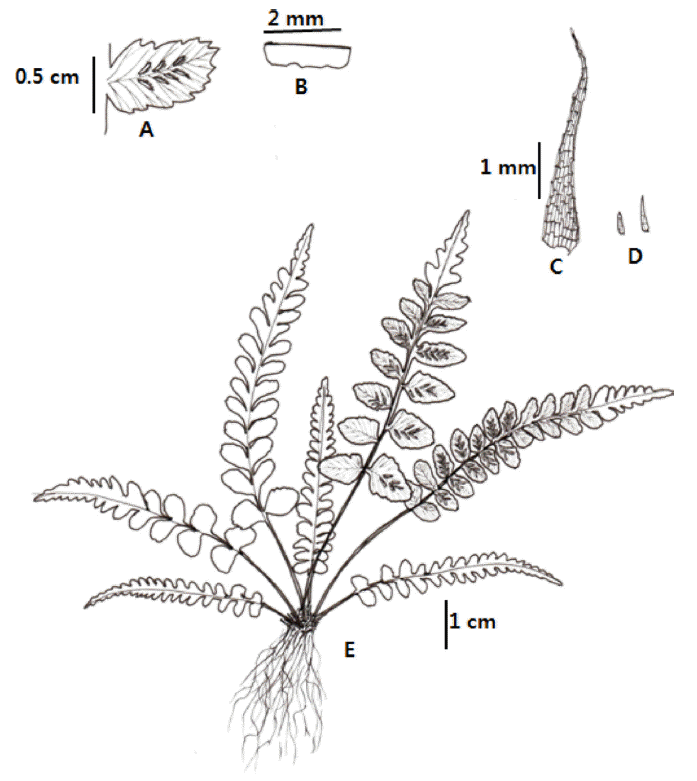
Korean name: Geo-mi-sa-cheol-go-sa-ri (거미사철고사리) TYPE: Korea. Seoul, Mt. Bukansan, elev. 178 m, 27 Aug. 2013, C.S. Lee & K. Lee 13082701 (holotype, EWH: isotype, KB) (Fig. 1)
Winter green herb, on rock or epiphytic, height 5−15 cm. Rhizomes short gathering, 1.2−1.5 mm diameter, scaly; scales 3−5 mm long, 0.3−0.5 mm wide, narrowly lanceolate, blackish, almost entire. Stipes clustered, 1−3 cm long, 1−1.2 mm wide, green, base densely scaly, with reduced scales like hair toward rachis or subglabrous; lamina 1−2-pinnatifid, 4−12 cm long, 1.2−2.5 cm wide, linear-lanceolate, the apex abruptly to gradually reduced, firmly herbaceous; rachis green, sparsely scaly like hair (0.8−1.0 mm long, 0.010−0.015 mm wide), narrowly winged in upper lamina; lateral pinnae 8−12 pairs, oblong, basal basiscopic margin serrate, apex a little acuminate, 0.7−1.4 cm long, 0.4−0.7 cm wide, short-petiolulate (1.0−1.5 mm long); ultimate segments linear lanceolate, 3−4 mm long, 1−2 mm wide on basal pinnae, margin short toothed, never overlap; veins in abaxial surface protruded, 1 or 2 times forked; indusium slightly lacerate; sori 1.5− 3.0 mm long; spores abortive.
Habitat: On the rock near mountain stream.
Distribution: Uiryeong-gil, Mt. Bukansan, Seoul, Korea
Note: Gross morphology of putative parental species, A. ruprechtii and A. pekinense, and their putative hybrid, A. × uiryeongse, are shown in Figs. 1, 2, and 6. Typical A. × uiryeongse has intermediate characters between the parents such as narrowly winged upper rachis, 1−2-pinnatifed lamina, shorter pinna stalk, shape of pinna and pinnule, and serration of indusium margin (Table 1, Figs. 1, 2, and 6). Asplenium × uiryeongse has some characters of A. ruprechtii, such as entire scale margin, no shorter base of lamina, and stipe length, as well as characters similar to A. pekinense, such as oblong pinna, needle shaped pinnule segment, and lacerated margin of indusium.
Hybrids are known to have morphological characters intermediate between their parents as well as combining characters from each of the parents, although sometimes hybrids display characters not found in either parent (Grant, 1981; Rieseberg and Ellstrand, 1993). The putative hybrid A. × uiryeongse displays all of these inheritance patterns of parental characters. Furthermore, this taxon is an allotriploid hybrid (3x) of both putative parental species, A. ruprechtii (2x) and A. pekinense (4x) based on Flow Cytometry data and chloroplast rbcL and rps4-trnS sequence data (Lee et al., unpubl.).
The new scientific name was assigned based on its habitat, which it occurs in Uiryeong-gil of Mt. Bukansan and the new local name ‘geo-mi-sa-cheol-go-sa-ri was given to the new taxon based on its parent’s names. This hybrid was found in Korea with Asplenium ruprechtii Sa. Kurata, Asplenium pekinense Hance, Cheilanthes argentea (S.G. Gmel) Kunze, Quercus mongolica Fisch. ex Ledeb., Robinia pseudoacacia. L, Rhododendron mucronulatum Turcz, and Deutzia uniflora Shirai in a forest of Mt. Bukansan, Seoul, Korea.
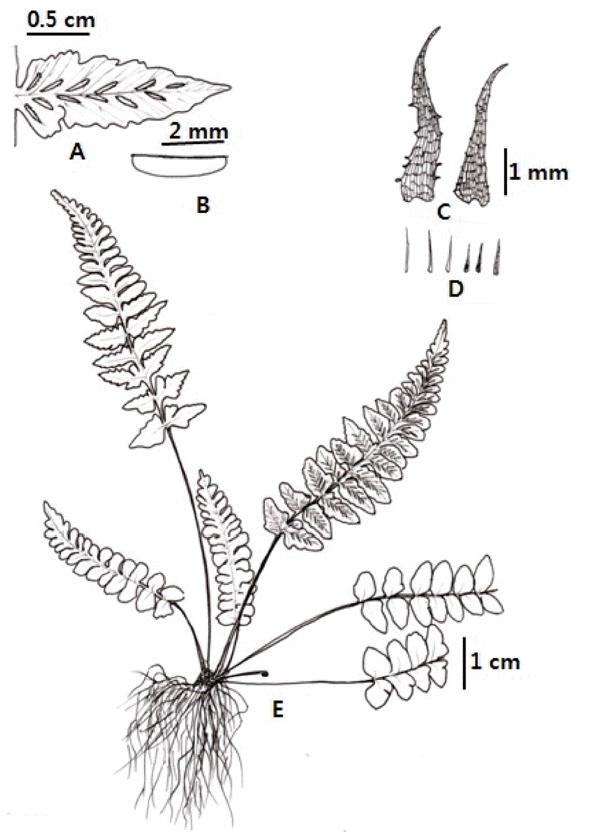
Korean name: San-kko-ri-go-sa-ri (산꼬리고사리)
TYPE: Korea. Geonggi-do, Yeoncheon-gun, Mt. Seongsan, elev. 468 m, 20 Sep. 2014, C.S. Lee & K. Lee 14092001 (holotype, EWH; isotype, KB, KH)
Winter green herb, on soils or rocks with many soils, height 5−12 cm. Rhizomes short gathering, 1.0−1.2 mm in diameter, scaly; scales 3-5 mm long, 0.5−0.7 mm wide, lanceolate, blackish brown, entire. Stipes clustered, 1−4 cm long, 1−1.2 mm wide, green, base densely scaly, with reduced scales like hair toward rachis; lamina 1-pinnate, 3−8 cm long, 1.3−2.0 cm wide, linear-lanceolate, the apex abruptly to gradually reduced, firmly herbaceous; rachis green, sparsely scaly like hair (0.3−1.0 mm long), winged in upper lamina; lateral pinnae 8−12 pairs, oblong or oblong-ovate, margin crenate, apex obtuse, 0.6−1.0 cm long, 0.5−0.7 cm wide, short-petiolulate (0.8−1.0 mm long), basal pinna largely serration two or three, never overlap; veins 2 or 3 times forked; indusium slightly lacerate; sori 1.5−3 mm long; spores abortive.
Habitat: On soils or rocks with rich soils under forests of high location of mountain.
Distribution: Mt. Seongsan, Yeoncheon-gun, Gyeonggi-do, Korea
Note: Gross morphology of putative parental species, A. ruprechtii, A. trichomanes and A. incisum, and their putative hybrid, A. × montanus, are shown in Figs. 3, 4, and 6. Typical A. × montanus has characters intermediate between the parents such as winged in upper rachis, 1-pinnate lamina, lamina quality, shape of pinna, and serration of indusium margin (Table 1, Figs. 3, 4, 6). Asplenium × montanus has some characters of A. ruprechtii, such as entire scale margin and shape of scale in stipe base, and no shorter base of lamina. It also has characters similar to A. trichomanes, such as oblong pinna, crenate pinna margin, and lacerated margin of indusium, as well as to A. incisum such as shape of basal pinna of lamina and soils or rocks with soil habitats.
The putative hybrid, A. × montanus displays all of these inheritance patterns of parental characters. Furthermore, this taxon (4x) occurred by reticulate evolution as a double hybrid between three putative parental species, A. ruprechtii (2x), A. trichomanes (4x), and A. incisum (2x) based on Flow Cytometry data and chloroplast rbcL and rps4-trnS sequence data (Lee et al., unpubl.).
The new scientific name was assigned based on its habitat, which it occurs in forest of mountain, and the new local name ‘san-kko-ri-go-sa-ri’ was given to the new taxon based on habitat, mountain. This hybrid found in Korea with Asplenium ruprechtii Sa. Kurata, Asplenium trichomanes L., Asplenium incisum Thunb., Quercus mongolica Fisch. ex Ledeb., Pinus densiflora Siebold & Zucc., and Rhododendron mucronulatum Turcz in a forest of Mt. Seongsan, Yeoncheon-gun, Gyeonggi-do, Korea. Even though A. × montanus is restricted to Yeoncheongun, its wide distribution and many individuals per population suggest that A. × montanus may be a stabilized hybrid derivative.
Asplenium × kitazawae Kurata & Hutoh
Voucher specimens: Korea. Daegu-si, Do-dong, elev. 120 m, 31 Aug. 2013, C.S. Lee & K. Lee 13083101 (EWH [2sheets]; 25 Sep. 2014, C.S. Lee & K. Lee 14092501 (KH [2 sheets], KB [2sheets])
Winter green herb, on rock or epiphytic, height 7−16 cm. Rhizomes short gathering, 1.2−1.5 mm in diameter, scaly; scales 4−5 mm long, 0.8−0.9 mm wide, lanceolate, blackish, entire. Stipes clustered, 1−4 cm long, 1−1.2 mm wide, green, the lowest base purplish, base densely scaly, with reduced scales like hair toward rachis; lamina 1−2-pinnatifid, 6−13 cm long, 1.8−3.5 cm wide, oblong-lanceolate or lanceolate, the apex abruptly to gradually reduced, firmly herbaceous; rachis green, sparsely scaly like hair (0.8−1.0 mm long), winged in upper lamina; lateral pinnae 8−14 pairs, oblong, ovate or deltoid-lanceolate, asymmetrical, basal pinnae equal or slightly shorter than next, basiscopic upper side auriculate, usually basal basiscopic side serrate, apex obtuse or shallowly acuminate, 0.7−2.0 cm long, 0.4−0.7 cm wide, short-petiolulate (1.0−1.5 mm long), margin crenate, some overlap; veins 1 or 2 times forked; indusium entire; sori 1.0−1.5 mm long; spores abortive.
Habitat: In forests of mountain.
Distribution: Do-dong, Daegu-si, Korea.
Note: The newly reported taxon in Korea, Asplenium × kitazawae, is known as a hybrid of A. ruprechtii and A. sarelii from Japan (Iwatsuki, 1995). Gross morphology of putative parental species, A. ruprechtii, A. sarelii, and their putative hybrid, A. × kitazawae, are shown in Figure 5 and 6. Typical A. × kitazawae has intermediate characters between the parents such as shape of scale in stipe base, winged in upper rachis, 1−2-pinnatifid lamina, lamina quality, shape and margin of pinna (Table 1, Figs. 5, 6). Asplenium × kitazawae has some characters of A. ruprechtii, such as narrow lamina, shorter stipe, and shorter indusium. It also has characters similar to A. sarelii such as serrated lamina, oblong or deltoid pinna, no gemma in apex of lamina.
The putative hybrid, A. × kitazawae displays all of these inheritance patterns of parental characters. Furthermore, this taxon is an allodiploid or allotetraploid hybrid (2x, 4x) of both putative parental species, A. ruprechtii (2x) and A. sareli (6x) based on Flow Cytometry data and chloroplast rbcL and rps4-trnS sequence data (Lee et al., unpubl.).
The local name was designated as ‘geo-mi-dol-dam-go-sa-ri’ from based on its parent name. It was found in Korea with Asplenium ruprechtii Sa. Kurata, Asplenium sarelii Hook., Platycladus orientalis (L.) Franco, Selaginella tamariscina (P. Beauv.) Spring, Pyrrosia linearifolia (Hook.) Ching, and Rhododendron mucronulatum Turcz in a forest of Dodong, Daegu-si, Korea.