UNDRUUL, MUNKHTULGA, BAASANMUNKH, OYUNTSETSEG, TSEGMED, SER-ODDAMBA, and CHOI: Two new alien records of Hordeum jubatum (Poaceae) and Tripleurospermum inodorum (Asteraceae) in Mongolia
Abstract
Hordeum jubatum L. and Tripleurospermum inodorum (L.) Sch. Bip. are currently recorded as non-native in the flora of Mongolia. Hordeum jubatum can easily be distinguished from other Mongolian Hordeum species by its glumes that are much (usually more than 10×) longer than the floret. Tripleurospermum inodorum differs from T. limosum (Maxim.) Pobed. by its annual or biennial plants without rhizomes, branched stems, and capitulae. The present study provides descriptions, a distribution map, taxonomic keys, taxonomic notes, and photographic illustrations of the two species based on newly collected samples.
Keywords: alien species, Hordeum, Tripleurospermum, flora of Mongolia, new record
INTRODUCTION
Alien vascular plant species are distributed worldwide ( Stohlgren et al., 2011; Seebens et al., 2021), with number of alien species constantly being discovered. The current knowledge of alien species of vascular plants in Mongolia is relatively low compared to that in Asian countries, owing to limited field surveys. Mongolian checklists of vascular plants, including both native and non-native species, have been published ( Grubov, 1982; Urgamal et al., 2014). The checklist of Mongolian vascular plants was recently updated based on the native species, comprising 3,041 taxa from 653 genera and 111 families ( Baasanmunkh et al., 2022). Approximately 60 non-native species have been recorded in Mongolia according to Baasanmunkh et al. (2022). Furthermore, Centaurea cyanus L. (Asteraceae), an alien species, was recently discovered in Mongolia ( Kechaykin et al., 2022). Poaceae is one of the world’s largest flowering plant families with 11,783 species and 789 accepted genera ( Soreng et al., 2022). They are annual or perennial herbs that have the following morphological characteristics—one-seeded indehiscent fruit, leaves that are parallel-veined and two-ranked, a ligule that commonly consists of flaps of tissue or hair, and a spikelet with 1–many ( Peterson, 2013). The structure of the ligule, along with the degree of sheath closure, has been indicated to be of great importance among the macromorphological characteristics ( Tzvelev, 1989). Hordeum L. is the smallest genus in the Poaceae family, comprising 34 taxa worldwide ( Blattner, 2004; Plants of the World Online, 2023). This genus belongs to the Triticeae tribe, which is particularly differentiated by its spike inflorescence compared to the panicle inflorescence frequently observed in other grass species, open leaf sheath with membranous ligules, and hairy top of the developing grain ( Blattner, 2018). Species of Hordeum are distributed throughout the Northern Hemisphere, southern South America, and South Africa ( Bothmer et al., 1995). A total of seven species of Hordeum have been recorded in Mongolia, including four native and three nonnative species ( Grubov, 1982; Baasanmunkh et al., 2022). Asteraceae is one of the mega-diverse families, with an estimated 25,000–35,000 species, 1701 accepted genera comprise 10% of all flowering plant species, which are widespread throughout the earth ( Mandel et al., 2019; Plants of the World Online, 2023). Tripleurospermum Sch. Bip. is a member of the Anthemideae tribe, the seventh-largest tribe of Asteraceae ( Oberprieler et al., 2007). This genus is primarily distributed in the Mediterranean Basin, including Europe, temperate Asia, and North Africa ( Oberprieler et al., 2007). The presence of coronate achenes with thick obtuse ribs is considered a valuable morphological trait for distinguishing Tripleurospermum taxa ( Schischkin and Bobrov, 2000). The genus Tripleurospermum comprises 41 species and is primarily distributed in the Northern Hemisphere (Plants of the World Online, 2023). Of these, only Tripleurospermum ambiguum (Ledeb.) Franch. & Saw. were observed in western Mongolia ( Baasanmunkh et al., 2022). This study discovered two alien species in Mongolian flora, Hordeum jubatum L. and Tripleurospermum inodorum (L.) Sch. Bip. and provides a distribution map, taxonomic keys, photo illustrations, and habitat information for each species.
MATERIALS AND METHODS
Field surveys were conducted in Ulaanbaatar, Mongolia, in 2023. Wild photographs of each species were also captured. The collected specimens were deposited in the herbarium of the National University of Mongolia (UBU) and official herbarium barcodes were provided. For the distribution map, point data for each species were obtained from two sources: (1) field survey collections in 2023 and (2) the “Flora of Mongolia” project on the iNaturalist platform, established in January 2019 ( https://www.inaturalist.org/projects/flora-of-Mongolia) ( iNaturalist, 2023). The general land cover of the selected sites was downloaded from the ESA World Cover ( Zanaga et al., 2022) to create a distribution map in ArcGIS ( Esri, 2012).
RESULTS AND DISCUSSION
A comprehensive investigation of the relevant literature and illustrations of T. inodorum ( Figs. 1, 3B) and H. jubatum ( Figs. 2, 3A) was conducted to ensure precise identification and complete morphological descriptions. A distribution map, taxonomic keys, photo illustrations, and habitat information were provided for each species. The taxonomic keys of new records of H. jubatum and T. inodorum were added to the representative genera of Grubov’s work ( Grubov, 1982). Both species are found in urban areas in Mongolia. During the field surveys, we collected the herbarium specimens for each species from a single site, respectively. While, other distribution records were obtained from the Flora of Mongolia project in iNaturalist platfrom ( https://www.inaturalist.org/projects/flora-of-mongolia) ( Fig. 4).
Key to the species of Tripleurospermum in Mongolia
1. Plants perennial with rhizome; stems unbranched, scattered pilose below capitula; capitula solitary, terminal, 3–4 cm in diameter; corona with broad obtuse teeth ····················· ···························································· T. ambiguum (native) 1. Plants annual or biennial, without rhizome; stems corymbosely branched in upper 1/2, glabrous; capitula several to many, in an apical corymb, 2–3 cm in diameter; corona short, subentire ·············· T. inodorum (non-native)
Tripleurospermum inodorum (L.) Sch. Bip., Tanaceteen: 32, 1834. Matricaria inodora L., Fl. Suec., ed. 2: 297, 1755; Chrysanthemum inodorum (L.) L., Sp. Pl., ed. 2.: 1253, 1763; Pyrethrum inodorum (L.) Moench, Methodus (Moench), 597, 1794; Chamomilla inodora (L.) K. Koch, Linnaea 17: 45, 1843; Rhytidospermum inodorum (L.) Sch. Bip. in P. B. Webb & S. Berthelot, Hist. Nat. Iles Canaries (Phytogr.) 3: 277, 1844; Chamaemelum inodorum (L.) Vis., Giorn. Bot. Ital. 1: 34, 1845 ( Fig. 1).
Herbs annual or biennial. Stems erect, striate, glabrous, corymbosely branched in upper half, 30–100 cm tall. Leaves sessile in middle of stem; leaf blade ovate-oblong or oblong, 4–8 cm long, 1.5–5 cm broad, both surfaces glabrous, irregularly tripinnate into linear acute segments; distal stem leaves similar, gradually smaller. Capitula several to many, in an apical corymb, 2–3 cm in diam., pedunculate; involucres hemispheric, 7–10 mm in diam.; phyllaries in 3 or 4 rows, margin narrowly white scarious, outer ones lanceolate, middle and inner ones oblong to oblanceolate. Ray florets white; ligules 10–20 mm long. Disk florets yellow, 5-lobed; lobes each with a resin sac. Achenes brown, triquetrous, ca. 2 mm, wrinkled, with 1 adaxial and 2 lateral thick whitish ribs, abaxially and apically with 2 distinct resin sacs.
Flowering and fruiting: June to September.
Distribution: Native to Europe to Siberia and Xinjiang (Plants of the World Online, 2023).
Habitats: Tripleurospermum inodorum grows in the tree cultivated and urban areas. Currently, four locations are known across Ulaanbaatar city ( Fig. 4).
Specimen examined: MONGOLIA. Khentei region: Ulaanbaatar city, Sukhbaatar district, Dambadarjaa, 47°59′04.8″N, 106°56′56.0″E, 1,390 m, a.s.l., 28 Jul 2023, B. Oyuntsetseg & D. Munkhtulga (UBU0033766-UBU0033768).
Taxonomic note: Tripleurospermum inodorum (L.) Sch. Bip. is morphologically similar to T. limosum (Maxim.) Pobed. in its erect, striate stem, without rhizome, phyllaries with white or pale brown scarious margins, and wrinkled achenes. However, the two species differ in their ray florets, coronas, and leaf blade characteristics. Tripleurospermum inodorum has ray floret ligules 10–20 mm long and a sub-entire corona. In contrast, T. limosum has ray floret ligules 4.5 mm and a corona 3-toothed. In terms of the leaf blade, T. inodorum has ovate-oblong or oblong, 4–8 × 1.5–5 cm, both surfaces glabrous, 2-pinnatisect, and ultimate segments narrowly linear. Whereas, T. limosum has oblanceolate oblong or oblong, 5.5–9.5 × 2.5– 3 cm, both surfaces glabrous, tripinnatisect; ultimate segments filiform.
Key to the species of Hordeum in Mongolia
1. Plants perennial; glumes subulate-setaceous, not lanceolate dilated at base.
2. Glumes much (usually more than 10×) longer than floret········································································ H. jubatum (non-native) 2. Glumes shorter to slightly longer than floret.
3. Lemma awns in medial spikelets up to 4 mm long; anthers in fertile spikelets 2.8–4.3 mm long ··································· ·························································································································································· H. brevisabulatum (native) 3. Lemma awns in medial (fertile) spikelets 5–12 mm long; anthers in fertile spikelets 0.7-2 mm long.
4. Stems with short-pilose nodes. Lemmas on outside studded with elongate spinules throughout surface; anthers 1.2– 2 mm long ··········································································································································· H. bogdanii (native) 4. Stems with glabrous nodes. Lemmas on the outside only in upper part with short spinules; anthers 0.7–1.2 mm long ·· ···························································································································································· H. roshevitzii (native)
1. Plants annual; glumes lanceolate dilated at base.
5. Spikes distichous: in each group of 3 spikelets only medial one bisexual and fertile, lateral ones sterile (underdeveloped or staminate); lemmas of medial spikelet with long awns ······································ H. vulgare subsp. distichon (non-native) 5. Spikes hexastichous: all 3 spikelets in each group fertile, with bisexual flower.
6. Lemmas at apex with long awn ······································································································· H. vulgare (non-native) 6. Lemmas at apex with tripartite appendage, its shorter lateral lobes deflexed downward, and large medial one spurlike, curved ··············································································································································· H. aegiceras (non-native)
Hordeum jubatum L., Sp. Pl. 1: 85, 1753. Elymus jubatus (L.) Link, Hort. Berol. 1: 19, 1827; Critesion jubatum (L.) Nevski, V. L. Komarov (ed.), Fl. URSS 2: 721, 1934.—TYPE. Canada (LINN, No. 103: 10 according to Baden and Bothmer, 2004) ( Fig. 2).
Common names: foxtail grass, squirrel-tail grass.
Herbs perennial. Culms 20–50 cm tall, 2–2.5 mm in diam., 3–5-noded, glabrous and smooth, erect or decumbent at base. Leaves narrowly linear, 6–12 cm long, 1.5–4 mm broad, scabrous on both sides, occasionally hairy on the upper surface, green, longer than the internodes, glabrous and smooth, or the lower ones sometimes hairy. Spikes 3–7 cm long, not taking into account the surrounding awns, and 4– 7 mm broad, usually somewhat nodding, light green or somewhat purple-tinged. Lateral spikelets: reduced to 1–3 spreading awns, rarely male. Central spikelet perfect; glumes spreading, awnlike, 2.5–6.5 cm, much longer than floret; lemma lanceolate, 5–7 mm, awn 2.5–8 cm long; palea equaling lemma. Grains (seeds) are oval-elliptic and smooth except for short hairs at the tip.
Flowering and fruiting: July to August.
Distribution: Native in Siberia, the Caucasus, northern China, and North America (Plants of the World Online, 2023). Alien in South America, Europe, Central Asia, and South Korea ( Baden and Bothmer, 1994; Kil et al., 2001).
Habitats: Hordeum jubatum mostly grows in roadsides, behind of builds and slabs of buildings in urban environments. Based on distribution map, this species has been quite widely distributed in Ulaanbaatar city ( Fig. 4).
Specimen examined: MONGOLIA. Khentei region: Ulaanbaatar city, Bayanzurkh district, Jukov, 47°55′03.4″N, 106°56′55.0″E, 1,300 m, a.s.l., 19 Jul 2023, Z. Tsegmed & D. Munkhtulga (UBU0033761-UBU0033765).
Taxonomic note: Hordeum jubatum is perennial, and the glumes are longer than the flowers (usually more than 10×) making it easily distinguishable from other Hordeum. The native range of this species is generally from Europe to North Africa and South and North America ( Britton and Brown, 1970). It is distributed as a rare, adventive weed in Western Siberia, Russia’s European region, and Asia. Hordeum jubatum has not been mentioned in previous floristic works in Mongolia (i.e., Grubov 1982; Urgamal et al., 2014), therefore, not native to Mongolia.
Conclusion: Considering the recently recorded occurrences of these two species, 62 species from 44 genera within nonnative taxa were identified in the flora of Mongolia. There is currently not much information available regarding alien plants in the country due to limited field surveys. Further revisions to the non-native taxon checklist are required to control for regional plant invasion. Future findings on species must be prioritized to ensure the effective control of invasive alien species. It is believed that this report will help in prioritizing notes on non-native species in Mongolia.
ACKNOWLEDGMENTS
This work was supported by the Korea National Arboretum (Grant Number: KNA1-1-26, 20-1) and Korea Basic Science Institute (Nantional Research Facilities and Equipment Center) grant funded by the Ministry of Education (Grant No. 2023R1A6C101B022).
Fig. 1.
Tripleurospermum inodorum in Ulaanbaatar, Mongolia. A, B. Habit. C. Flowers. D. Leaves. E. Ray floret. F. Involucral bracts. 
Fig. 2.
Hordeum jubatum in Ulaanbaatar, Mongolia. A, B. Habit. C, D. Portion of the spike. E, G. Spikelets. F, H. Sheath, ligule, and blade. 
Fig. 3.
Voucher specimens. A. Hordeum jubatum. B. Tripleurospermum inodorum. 
Fig. 4.
Distribution map of Hordeum jubatum (black dot), and Tripleurospermum inodorum (red square) in Ulaanbaatar, Mongolia 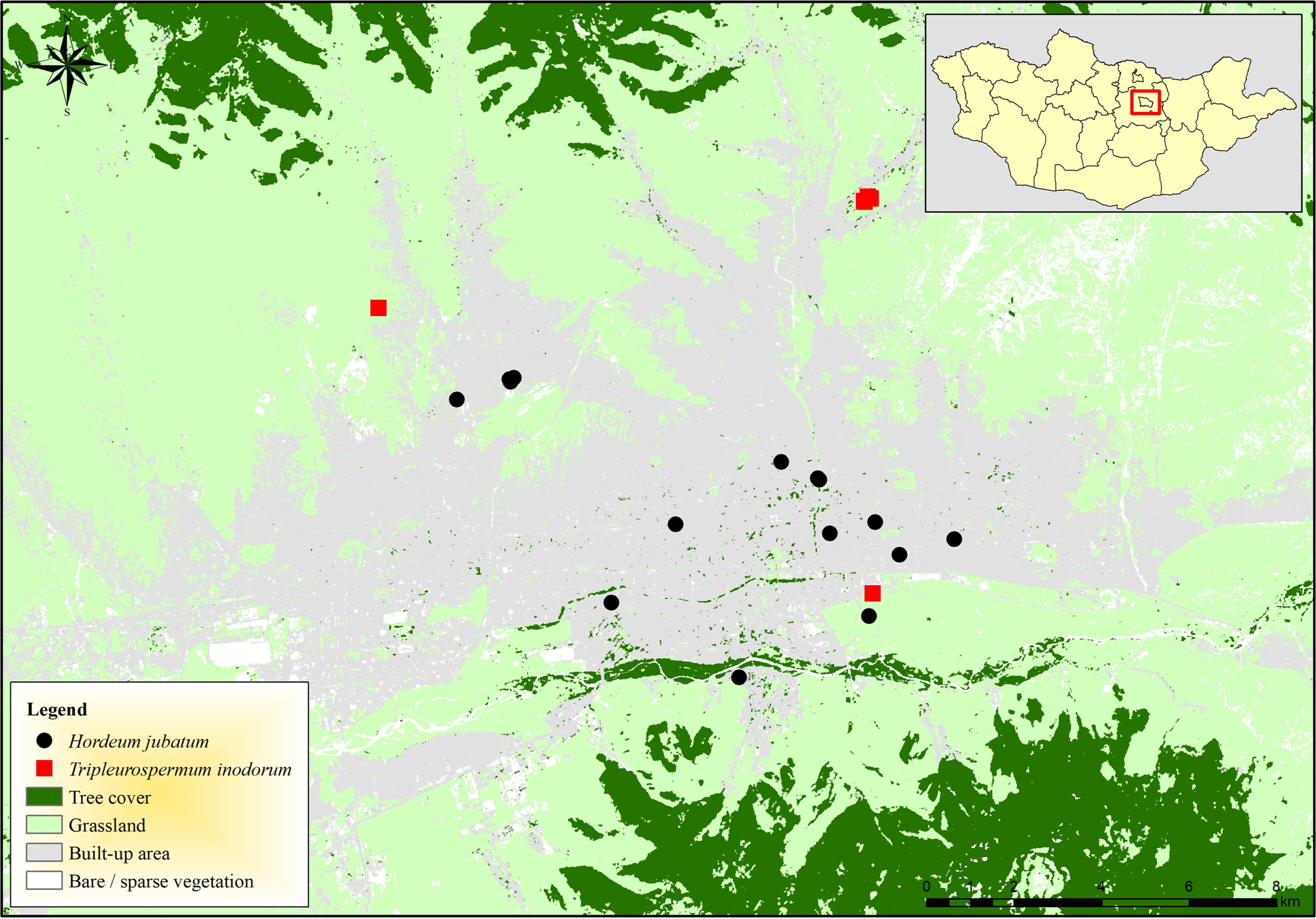
LITERATURE CITED
Baasanmunkh, S. Urgamal, M. Oyuntsetseg, B. Sukhorukov, A. P. Tsegmed, Z. Son, D. C. Erst, A. Oyundelger, K. Kechaykin, A. A. Norris, J. Kosachev, P. Ma, J.-S. Chang, K. S and Choi, H. J. 2022. Flora of Mongolia: annotated checklist of native vascular plants. PhytoKeys 192: 63-169.     Baden, C and von. Bothmer, R. 1994. A taxonomic revision of Hordeum sect. Cristerion
. Nordic Journal of Botany 14: 114-136.
Blattner, F.R. 2004. Phylogenetic analysis of Hordeum (Poaceae) as inferred by nuclear rDNA ITS sequences. Molecular Phylogenetics and Evolution 33: 289-299. Blattner, F. R. 2018. Taxonomy of the genus Hordeum and Barley ( Hordeum vulgare). The Barley Genome. Stein, N. Muehlbauer, G (eds.), Springer, Cham. 11-23. Bothmer, R. von. Jacobsen, N. Baden, C. Jorgensen, R. B and Linde-Laursen, I. 1995. An Ecogeographical Study of the Genus Hordeum. 2nd ed. International Plant Genetic Resources Institute, Rome. 129 pp.
Britton, N. L and Brown, A. 1970. An Illustrated Flora of the Northern United States and Canada, Vol. 1. Dover Publications, New York. 287-288.
Esri. 2012. ArcGIS Desktop: Release 10.2. Environmental Systems Research Institute, Redlands, CA, USA.
Grubov, V. I. 1982. Key to the Vascular Plants of Mongolia. Nauka, Leningrad. 433 (in Russian).
Kechaykin, A. A. Shmakov, A. I. Batkin, A. A. Gundegmaa, V. Baasanmunkh, S. Oyuntsetseg, B. Choi, H. J. Gregor, T. Paule, J. Shaulo, D. N. Korolyuk, A. Y. Zibseev, E. G. Sinitsyna, T. A and Smirnov, S. 2022. New findings in the flora of Mongolia, part 2. Turczaninowia 25: 105-123. Kil, J.-H. Suh, M.-H and Park, S.-H. 2001. Unrecorded and naturalized plants in Korea. Korean Journal of Plant Taxonomy 31: 375-382.
Oberprieler, C. Himmelreich, S and Vogt, R. 2007. A new subtribal classification of the tribe Anthemideae (Compositae). Willdenowia 37: 89-114. Schischkin, B. K and Bobrov, E. G. 2000. Flora of the USSR, Vol. 26. Compositae, Tribes Anthemideae, Senecioneae, and Calenduleae. Smithsonian Institution Libraries, Washington, D.C.. 892 pp.
Seebens, H. Blackburn, T. M. Hulme, P. E. van Kleunen, M. Liebhold, A. M. Orlova-Bieñkowskaja, M. Pyšek, P. Schindler, S and Essl, F. 2021. Around the world in 500 years: Interregional spread of alien species over recent centuries. Global Ecology and Biogeography 30: 1621-1632. Soreng, R. J. Peterson, P. M. Zuloaga, F. O. Romaschenko, K. Clark, L. G. Teisher, J. K. Gillespie, L. J. Barberá, P. Welker, C. A. D. Kellogg, E. A. Li, D.-Z and Davidse, G. 2022. A worldwide phylogenetic classification of the Poaceae (Gramineae) III: An update. Journal of Systematics and Evolution 60: 476-521. Stohlgren, T. J. Pyšek, P. Kartesz, J. Nishino, M. Pauchard, A. Winter, M. Pino, J. Richardson, D. M. Wilson, J. R. U. Murray, B. R. Phillips, M. L. Ming-yang, L. Celesti-Grapow, L and Font, X. 2011. Widespread plant species: Natives versus aliens in our changing world. Biological Invasions 13: 1931-1944. Tzvelev, N. N. 1989. The system of grasses (Poaceae) and their evolution. Botanical Review 55: 141-203. Urgamal, M. Oyuntsetseg, B. Nyambayar, D and Dulamsuren, C. 2014. Conspectus of the Vascular Plants of Mongolia. Admon Printing, Ulaanbaatar. 333 pp.
Zanaga, D. Van De Kerchove, R. Daems, D. De Keersmaecker, W. Brockmann, C. Kirches, G. Wevers, J. Cartus, O. Santoro, M. Fritz, S. Lesiv, M. Herold, M. Tsendbazar, N.-E. Xu, P. Ramoino, F and Arino, O. 2022. ESA* European Space Agency, WorldCover, version v200. Zenodo
|
|











