INTRODUCTION
Throughout the world, climate change, pollution, and natural destruction occur as a consequence of human actions, such as agricultural production, fish harvesting, and the exploitation of natural resources. As a result, various species have become extinct. It is estimated that approximately 25% of animal and plant species encounter distinction in the world, and the average abundance of native species has decreased by at least 20% (Díaz et al., 2019). The loss of biodiversity has caused a decline in the resilience of the ecosystem.
To prevent species from becoming extinct, efforts to designate species as endangered and protect their habitats are needed. As public awareness of the importance of endangered species and biodiversity has increased, the Korean government has been participating in the Convention on Biological Diversity (CBD) since 1995. The government is also making efforts to conserve endangered species in Korea by enacting the Biodiversity Conservation and Use Act and announcing the Comprehensive Plan for the Conservation of Endangered Wildlife.
Globally, practical guidelines for species conservation have been published by international organizations such as the International Union for the Conservation of Nature (IUCN). According to the IUCN/SSC (2013), the act of an intentional organism transferred from one site for release in another to conserve the species is defined as conservation translocation. There are four categories of conservation translocation depending on the geographical range of the translocation and the purpose of the translocation. Among them, when the translocation occurs outside of the indigenous range when, for instance, the indigenous habitat becomes threatened and unsustainable, this type is categorized as assisted colonization. Assisted colonization is considered to be a good method for species conservation under climate change (Gallagher et al., 2015; Ferrarini et al., 2016).
The genus Iris L. consists of 225 species in the family Iridaceae, and is widely distributed in temperate zones of the Northern Hemisphere (Sim, 2007). Among the species of Iris, 13 species are reported to occur in the Korean peninsula and a number of rare plants, such as I. dichotoma, I. koreana, and I. odaesanensis, have been included of various studies (Sim, 2007; Jang and Lee, 2022).
Iris dichotoma Pall. (Iridacea), an endangered species in Korea, is a perennial plant distributed in north-west part of islands in Korea, such as Daecheong-do, Baengnyeong-do, and Taean (Chang et al., 2016; Ministry of Land Infrastructure and Transport, 2017). It usually grows in rocky cracks and grasslands at low elevations and has the trait of a floral circadian clock, opening around 3–4 pm and then closing before 10 pm. The dominant threats to I. dichotoma populations are known as plant collection by people, grazing by livestock in pastures, and a restricted habitat range (Korea National Park Research Institute, 2018).
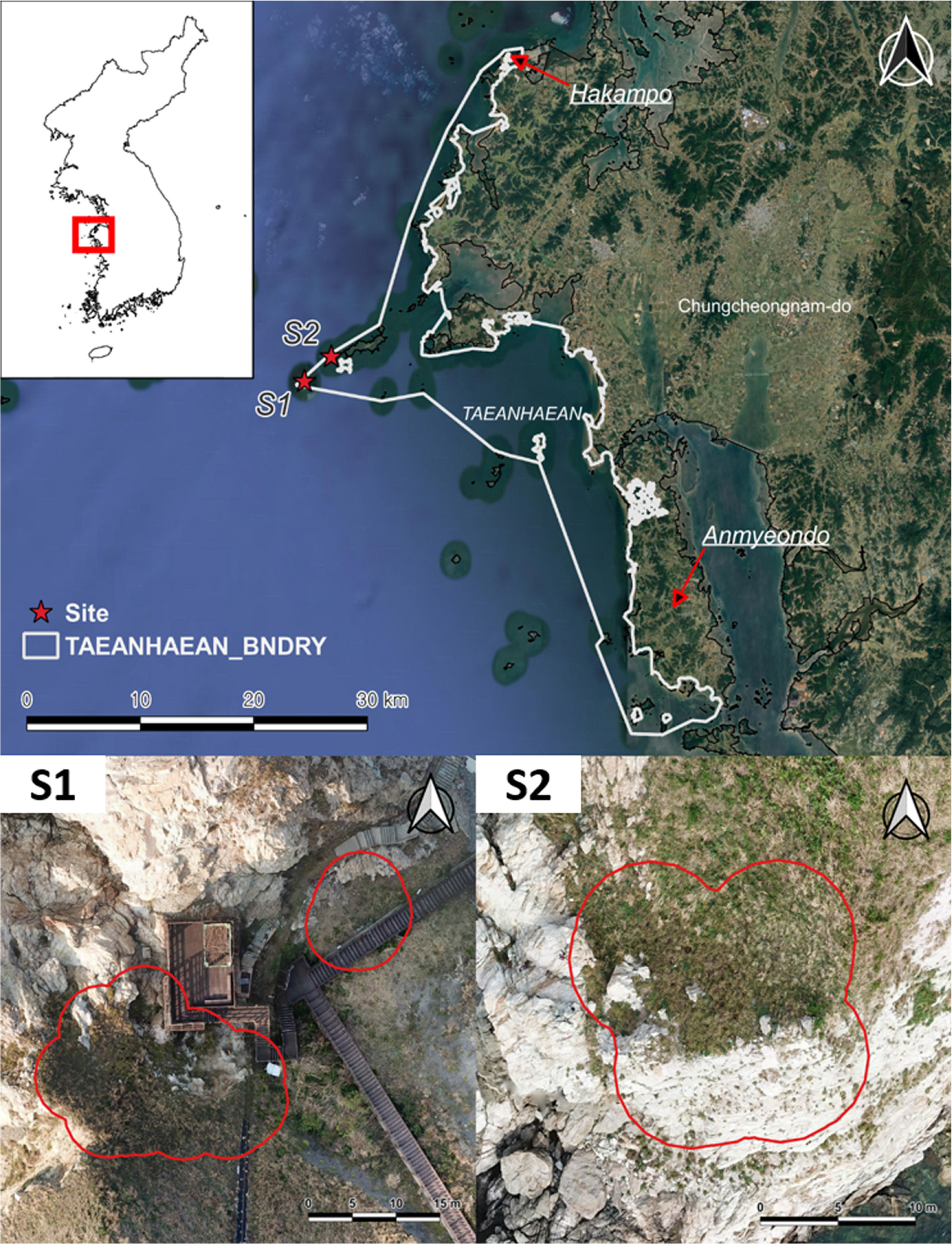
Since 1987, Korean national parks have been designated as locations at which to conserve nature resources in Korea. Among the three habitat sites of I. dichotoma, a site in Taean, found in 2013, is known as Taeanhaean National Park, one of Korea’s national parks. Thus, it was expected that the dominant threats to I. dichotoma may not arise at this site. However, the site was located near the coastline and is under the threat of ground weathering. Hence, a sustainable I. dichotoma habitat needed to be realized through assisted colonization in Taeanhaean National Park, near the indigenous habitat of this plant.
Previous studies of I. dichotoma included phylogenetic analyses (Park et al., 2002; Sim et al., 2002; Park et al., 2006; Lee and Park, 2013; Korea National Park Research Institute, 2018), callus induction and plant regeneration (Bae et al., 2012), seed germination (Lee et al., 2015; Park et al., 2021) and works on habitat characteristics and distribution range modeling (Korea National Park Research Institute, 2018), pollination studies (Liu and Fan, 2019;Liu et al., 2020), yearly seed weight determinations (Amartuvshin and Ochgerel, 2016) and possible medical uses (Wei et al., 2012). However, due to the rarity of the species and its limited habitat range, studies of the growth of I. dichotoma populations in the wild are rare, especially for individuals after conservation translocation.
The aim of this study is to describe changes in the annual growth of I. dichotoma and population density levels after conservation translocation. The following hypotheses were tested: (1) A survival and growth of cultivated I. dichotoma will be changed after transplanting in the wild and (2) the transplanted I. dichotoma will become similar to the nearby natural I. dichotoma gradually when the transplanted site is adequate for the species. To test these hypotheses, the following factors were measured: annual survival, population density, leaf bundle width (LBW), leaf bundle length (LBL), pedicel length, and flowering rate for three years.
MATERIALS AND METHODS
Preparation of Iris dichotoma
In 2013, two ovaries of natural I. dichotoma were collected in Taeanhaean National Park (Fig. 1). The seeds from the two ovaries were put into separate plastic pots filled with a 1:1 mixture of growing media and stone to imitate the soils in the natural habitat, and they were moved to a greenhouse bed. From 2015 to 2019, 100 pots of I. dichotoma were prepared gradually by repeated seed germination and root separation for this study.
Study site
To establish a sustainable habitat for the endangered I. dichotoma, a site without the threats in the original habitat site, such as ground weathering, was required. Two study sites were selected on islands that are part of Taeanhaean National Park (S1, S2) (Fig. 2). S1 was located on a slope opposite the slope of the natural I. dichotoma population (i.e., on the same island) and had an environmental condition similar to that of the natural habitat, and S2 was situated on another uninhabited island nearby. The vegetation types, geology, geography, wind directions, and the amounts of sunshine were similar between the sites (Table 1). The expected areas where I. dichotoma could disperse were 250 m2 for S1 and 110 m2 for S2.
Translocation and monitoring
In August 2019, the LBW (cm), LBL (cm), and pedicel length (cm) of natural I. dichotoma were surveyed in natural habitats in Taeanhaean National Park. The number of flowering individuals and clonal propagation were assessed as well. The pedicel length and flowering could not be measured when the individuals did not achieve flowering. In September of 2019, to monitor changes in the growth of I. dichotoma before and after translocation, the LBW (cm), LBL (cm), and pedicel length (cm) of cultivated I. dichotoma were measured with measuring tape in the greenhouse bed (Fig. 3). Flowering and clonal propagation were also checked. In October 2019, 70 pots of cultivated I. dichotoma (97 individuals in total due to clonal propagation occurring in each pot) were transplanted in S1 and 30 pots of I. dichotoma (36 individuals) were transplanted in S2. The number of pots at each site depended on the expected area for dispersal. Although clonal propagation also occurred in 2019, these individuals in pots were planted together at a set location. As I. dichotoma grows on rocky cracks along slopes and the cracks were randomly positioned, the planting distance could not be equal.
After translocation, the survival and number of instances of clonal propagation of I. dichotoma were checked every year (2020–2022). The flowering rate (%, [the number of flowering individuals/individuals that survived in the year × 100]) and clonal propagation increase (%, [the number of individuals in the year/the number of transplanted individuals in the beginning × 100]) were calculated. The population density (individual/m2, [the number of individuals that survived in the year/the area of the study site]) was also determined. The LBW, LBL, and pedicel length were also measured during the flowering season (August–September). As light competition and interruptions by other plants were possible threats at each site, other plants growing close to the transplanted I. dichotoma were removed during the monitoring period.
Statistical analysis
A repeated-measures ANOVA was conducted to monitor the effects of time and the sites on the growth (LBW, LBL, and pedicel length) of I. dichotoma at the two sites. In this analysis, the pedicel length was considered to be zero when an I. dichotoma individual did not flower during the year. Then, a generalized linear model (glm) and an lsmeans comparison test were used to compare the growth of cultivated I. dichotoma, transplanted I. dichotoma and natural I. dichotoma statistically. All statistical analyses were carried out using the SAS 9.4 software package (SAS Institute Inc., Cary, NC, USA).
RESULTS
Survival and population density after translocation
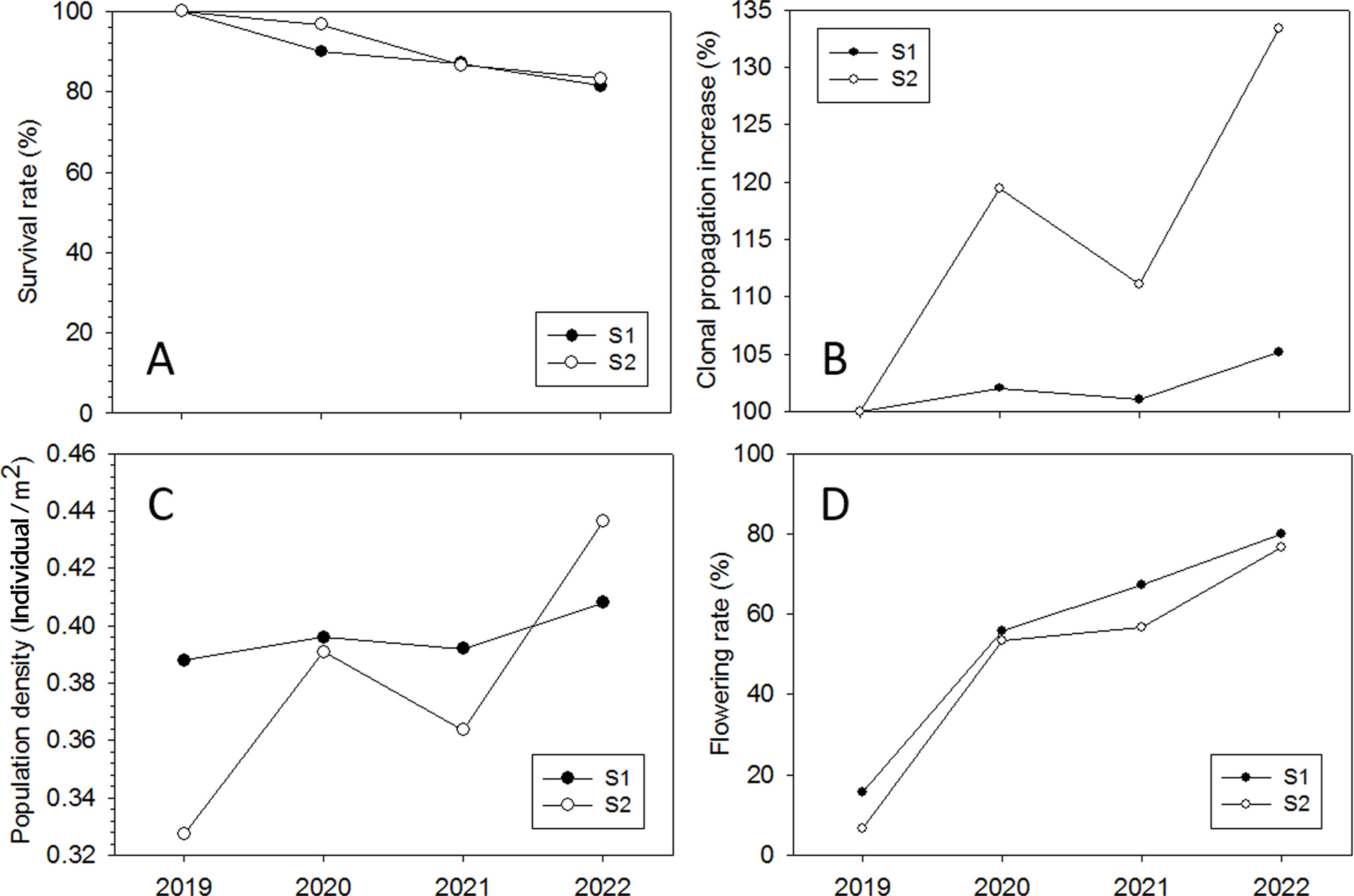
In 2022, the mean survival rate (%) of I. dichotoma after translocation was 82.4%. The survival rate (%) was similar at each study site (81.4% in S1 and 83.3% in S2) (Fig. 4A). The clonal propagation rate (%) increased every year (Fig. 4B). Consequently, the population density (individual/m2) increased from 0.39 to 0.41 in S1 and from 0.33 to 0.44 in S2 (Fig. 4C). The mean flowering rate (%) was only 11.2 in the cultivated I. dichotoma in 2019, but it reached 54.5 in 2020 and 78.33 in 2022 after transplanting (Fig. 4D).
Growth after translocation
The LBW and LBL outcomes of I. dichotoma were statistically different at the time of the measurements (year); however, the effects of the site were not significant (Table 2). The highest mean LBW and LBL values were found in cultivated I. dichotoma in 2019 (p < 0.05) (Table 3). Neither time nor site significantly affected the pedicel length (p > 0.05).
Comparison of natural, cultivated, and transplanted I. dichotoma populations
The average population density (individual/m2) was 0.52 for natural I. dichotoma, 0.36 when cultivated I. dichotoma was transplanted in the field, and 0.42 for the transplanted I. dichotoma in 2022. The mean flowering rate (%) for natural I. dichotoma was 75, similar to that of transplanted I. dichotoma in 2022, for which the rate was 78. Cultivated I. dichotoma showed the highest LBW, LBL, and pedicel length outcomes (Table 3). In contrast, the LBW, LBL, and pedicel length were statistically similar between the natural population and the transplanted population (p < 0.05).
DISCUSSION
This study was conducted to evaluate changes in I. dichotoma, an endangered species in Korea, after conservation translocation. Seeds from its natural habitat in Taeanhaean National Park were collected, and 100 pots of cultivated I. dichotoma in a greenhouse bed were prepared for this study. The cultivated I. dichotoma pots were transplanted at the two study sites in 2019. The annual survival and growth rates of the transplanted I. dichotoma were surveyed for three years.
The results of this study showed that although the survival rate of transplanted I. dichotoma decreased slightly, the population density and flowering rate improved overall. On the basis of these results, the study sites in this study were deemed to have adequate environmental conditions for I. dichotoma, and the newly transplanted I. dichotoma population will likely adapt well. According to research on the habitat of I. dichotoma (Korea National Park Research Institute, 2018), important factors affecting the growth of I. dichotoma could be sunshine and the presence of the other plants competing for light with I. dichotoma. The increase in clonal propagation was accord with results from Liu et al. (2019), who showed that I. dichotoma reproduction involves more clonal propagation than seed propagation.
The I. dichotoma cultivated in 2019 presented the greater LBW and LBL values than the transplanted I. dichotoma; however, flowering by the cultivated I. dichotoma was rarely achieved. These results show that the growth of I. dichotoma could be improved in a greenhouse bed, though reproduction rates may fall as well. Thus, I. dichotoma may allocate more energy to growth compared to reproduction in an environmental condition where threats such as light competition and harsh temperature changes rarely occur and survival is assured. Moreover, after I. dichotoma was transplanted, the growth of the cultivated I. dichotoma changed. It was clear that the annual size became similar to those of natural I. dichotoma nearby. The results may have been caused by the similar environmental conditions, such as precipitation levels and sea breezes, which they shared for a year. These types of changes in the phenotype of perennial plants depending on various environmental conditions were also found in other studies (Kozlowski and Wiegert, 1986; Moriuchi and Winn, 2005).
This study provides baseline data for the growth of endangered I. dichotoma in nature and in greenhouse beds. The results demonstrated the possibility that the phenotype of I. dichotoma can change due to environmental conditions after conservation translocation. At the study sites, the specific threats that other natural I. dichotoma populations confront, such as ground weathering and pasture feeding, could not occur, however, the threats of light competition from other plants (e.g., Miscanthus sinensis) still remain. Consequently, this conservation translocation study is considered to be successful, and monitoring and conservation efforts are likely still necessary for the continued conservation of the endangered I. dichotoma in the future.