Stachyuraceae J. G. Agardh, a family endemic to East Asia, is mainly distributed from the Himalayas across subtropical China to Taiwan Island and the Japan Archipelago (Feng et al., 2020). This family is comprised of a single genus, Stachyurus Siebold & Zucc. with approximately 11 species, and consists of shrubs and small trees (Keiji, 1959; Feng et al., 2020). Su et al. (2020) redefined seven species of Stachyurus following an integrative species concept and revised the taxonomy of Stachyuraceae based on previous reports and specimens, including the type specimens.
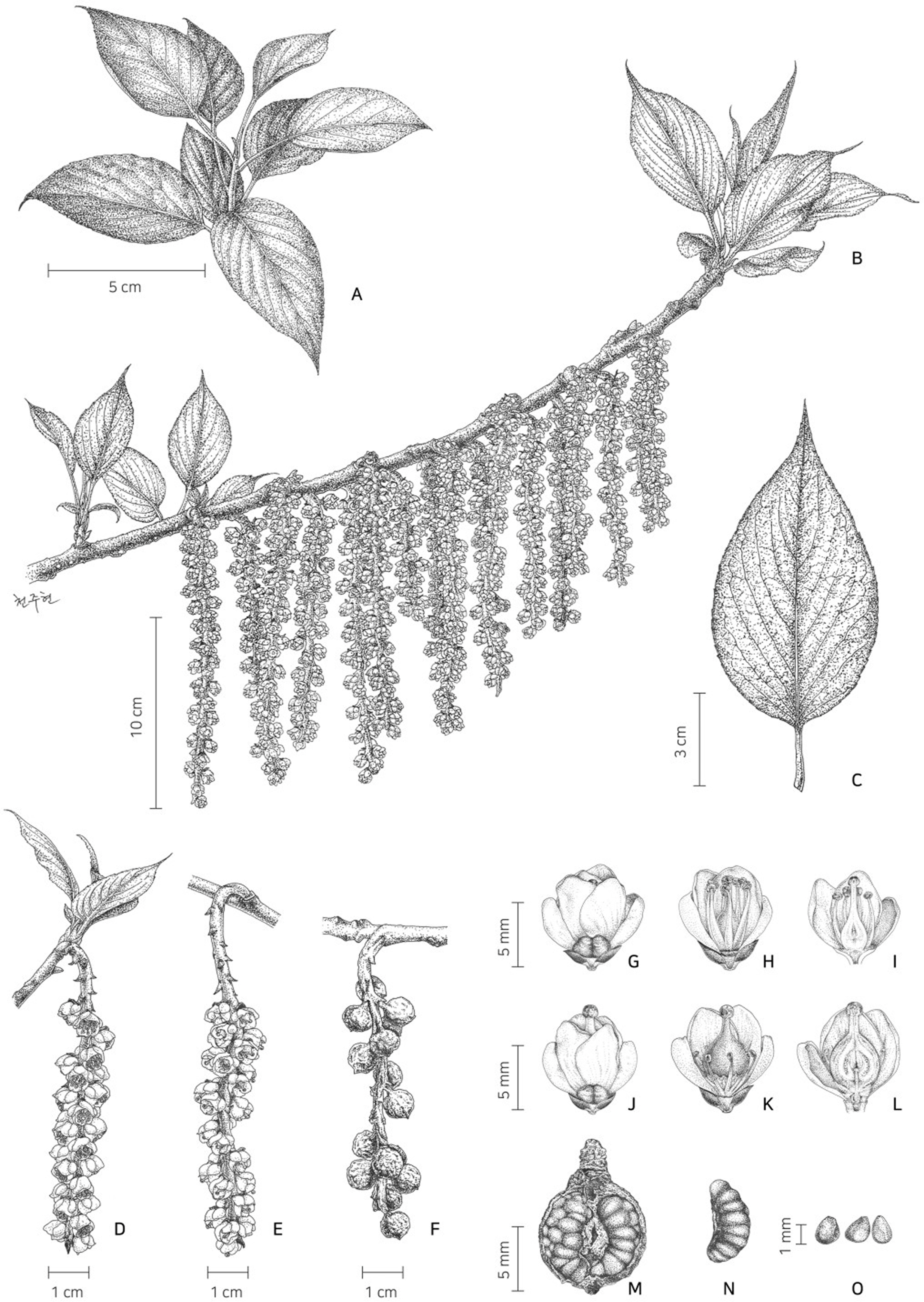
Stachyurus praecox Siebold & Zucc. has been regarded as an endemic species of Japan (Kitamura and Murata, 1994; Kato and Ebihara, 2011). This woody species is a pioneer shrub common in forest edges in the warm temperate zone in Japan (Abe, 2007). It is a small deciduous tree with single, alternate, and petiolate leaves (Keiji, 1959). Inflorescence is racemose, pendulous, located on axils of the previous year’s branches, and 3–10 cm in length. The flowers are hermaphrodite or sometimes plant dioecious, with four each of sepals and petals (Abe, 2007). Fruit is a berry with a coriaceous pericarp that are broadly ellipsoid to ovoid, 0.7–1.2 cm in diameter, and green and brownish yellow when mature (Feng et al., 2020; Su et al., 2020).
Recently, a natural population of the species was discovered on the uninhabited island of Wando-gun, Jollanam-do in Korea. This is the first report of S. praecox in a natural habitat in Korea. We provide the morphological characteristics, illustrations, photographs, and a distribution map of this woody species.
Materials and Methods
DNA extraction
Two accessions of S. praecox collected in Wando-gun were included in phylogenetic analysis. Voucher specimens (C. J. Oh OCJ80118 and OCJ80119) were deposited in the herbarium of Korea National Arboretum (KH). Specific informatin of the specimens are described in the Specimens examined section below. Total DNA from leaves was isolated using Exgene Plant SV (GeneAll Co., Seoul, Korea) following the manufacturer’s protocol. Isolated DNA was stored at −20ºC.
Primer design and polymerase chain reaction
PCR was performed using the internal transcribed spacer (ITS) 1 and ITS4 primers (White et al., 1990) for amplification of the ribosomal ITS regions. The PCR reactions were conducted with a final reaction volume of 30 μL containing EmeraldAmp GT PCR Master Mix (Takara, Tokyo, Japan), each primer (10 pM) and template DNA (50–100 ng/μL).
The PCR reactions were carried out with an initial denaturation for 5 min at 94ºC, followed by 35 cycles of denaturation for 30 s at 96ºC, annealing for 30 s at 57ºC, extension for 1 min at 72ºC, and with a final extension for 10 min at 72ºC. PCR products were electrophoresed on a 1.5% agarose gel, and then the DNA band was confirmed using UV transilluminator. Amplified PCR products were purified using AccuPrep PCR Purification Kit (Bioneer Co., Daejeon, Korea), and sequenced using the same primers at Macrogen Co., Seoul, Korea
Phylogenetic analysis
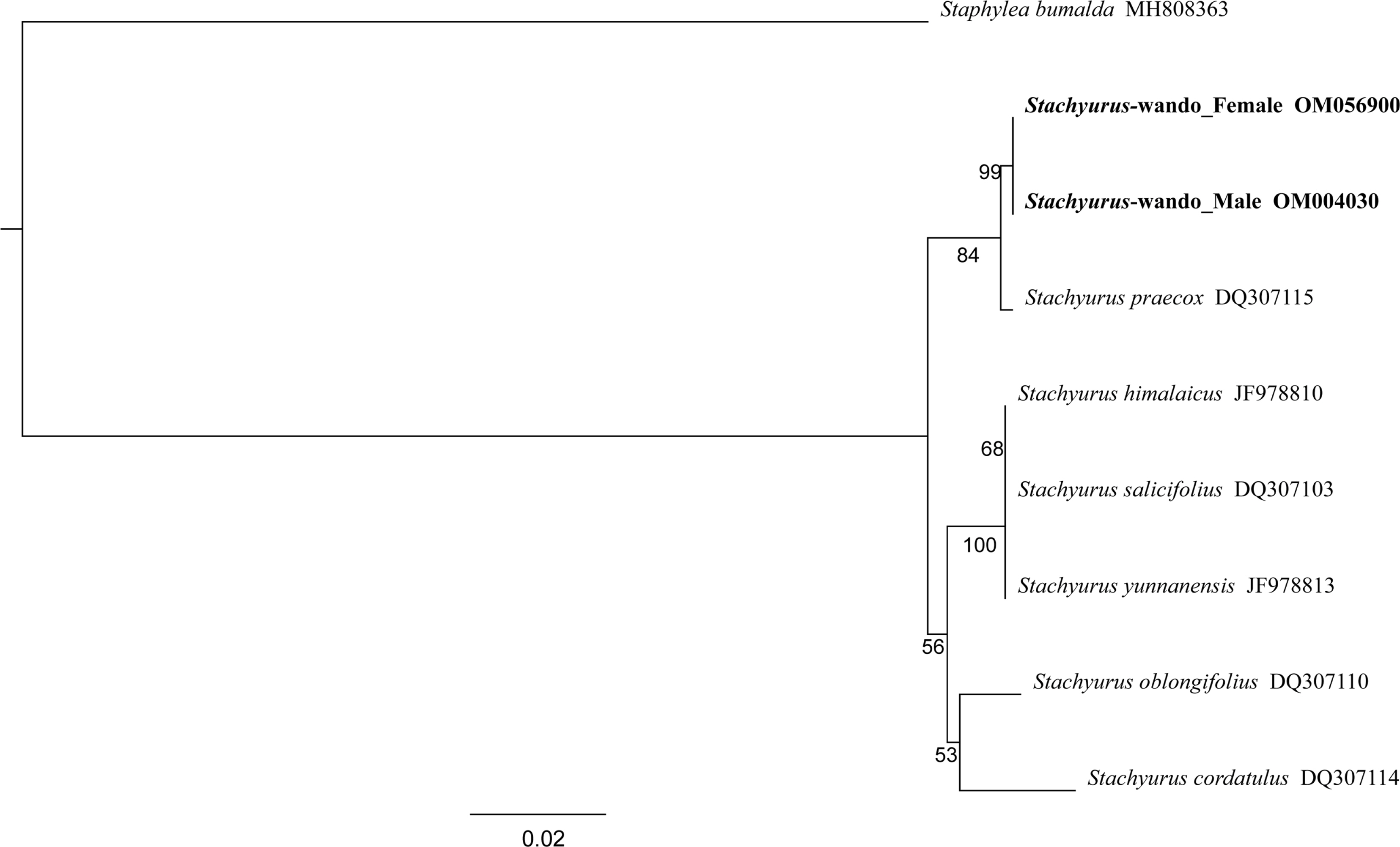
To determine the phylogenetic position of newly collected plants in this study, we conducted maximum likelihood analysis using the nucleotide sequences of the ITS regions of six species of Stachyurus. Also, for phylogenetic analysis, Staphylea bumalda DC. (GenBank accession number: MH808363) was included as an outgroup. The newly obtained ITS sequences were edited by Geneious Prime (Kearse et al., 2012), and multiple sequence alignment was conducted by using the MAFFT program (Katoh, 2013). Maximum Likelihood analysis of the ITS data was performed in RAxML 8.2.11 (Stamatakis, 2006) with the following settings. Nucleotide model was used GTR-GAMMA; algorithm was used Rapid Bootstrapping and search for best-scoring maximum likelihood tree; the number of starting trees or bootstrap replicates were performed 100 times; and parsimony random seed was performed.
Results and Discussion
To determine whether the samples of Stachyurus collected from Wando-gun, Jeollanam-do are the same taxon as S. praexox, Maximum likelihood analysis based on the ITS dataset of regions of six taxa of Stachyurus was performed. The results suggested two accessions representing male and female plants collected on Wando-gun, denoted as “Stachyurus-wando_ Male” and “Stachyurus-wando_Female” formed a clade with S. praexox with strong bootstrap support (Fig. 1). Therefore, Stachyurus collected from Wando-gun, Jollanam-do in the present study was considered to be in the same taxon as S. praexox distributed in Japan. This is the first report not only of Stachyurus praecox in Korea but of Stachyuraceae in the Korean flora. We suggest Wan-do-sul-kkot-na-mu-gua (완도술꽃나무과) for the Korean name of the family.
Taxonomic Treatment
Stachyurus praecox
Siebold & Zucc., Fl. Jap. (Siebold) 1: 43, 1836.
Stachyurus praecox var. rotundifolius Tuyama, J. Jap. Bot. 16: 504, 1940; S. praecox f. rotundifolius (Tuyama) Tuyama ex H. Hara, Enum. Sperm. Jap. 3. 229, 1954.
Stachyurus praecox var. microphyllus Nakai Bull. Nat. Sci. Mus. Tokyo no. 27: 31, 1949; S. praecox f. microphyllus (Nakai) H. Hara, Enum. Sperm. Jap. 3. 229, 1954.
Stachyurus praecox var. parviflous Makino ex H. Hara, Enum. Sperm. Jap. 3. 229, 1954.
Stachyurus praecox f. marginatus Hiyama, J. Jap. Bot. 36: 127, 1954.
Korean name: Wan-do-sul-kkot-na-mu (완도술꽃나무).
Deciduous usually small trees, stems 3–9 m tall, ca. 5–10 cm in diam., bark dull purplish-brown. Leaves alternate, petiole 1–3 cm long; blade chartaceous, usually elliptic to ovoid, 12–19 × 5.5–8 cm, apex acute to acuminate, base rounded to truncate, margins serrate, lower surfaces glabrous or sometimes pubescent on or along veins, lateral veins 5–7 pairs. Inflorescences racemose, pendulous, on axils of previous year’s branches, 10–18 cm long (usually shorter in female); flowers functionally dioecious (but morphologically hermaphrodite), campanulate, 7–9 mm long; pedicel less than 0.5 mm long; bract triangular, 1.5–2 mm long; sepals 4; petals 4, greenish-yellow in pistillate flowers; stamens 8, slightly shorter than petals, rudimentary in female flowers; pistil 1, slightly longer than stamens in staminate flowers; ovary narrowly ovoid; style shorter than ovary. Fruit berry, broadly ellipsoid to narrowly ovoid, 8–10 mm in diam., green, brownish-yellow when mature. Seeds 80–105 in a fruit, 1.5–3 mm long, 0.9–1.6 mm in diam., yellowish red (Figs. 2, 3).
Flowering: March to April.
Distribution: Korea (Wando-gun), Japan (Hokkaido, Honshu, Shikoku, Kyushu).
Specimens examined: KOREA. Jeollanam-do: Wando-gun, Saengil-myeon, Bongsun-ri, 4 Apr 2018, C. J. Oh OCJ80118, OCJ80119 (KH); 26 Mar 2021, C. J. Oh OCJ210088, OCJ210089, OCJ210090 (KH).
Stachyurus praecox was newly recorded from an inhabited island of Saengil-myeon, Wando-gun, Jeollanam-do in Korea (Fig. 4). Over 200 trees were distributed along the coastline on the island. The tallest tree was 8 m high and 23 cm in basal diameter. Most of the trees had two to 6 branches (Fig. 2).
Stachyurus praecox, distributed in Wando-gun, Jeollanam-do in Korea, is presumed to have migrated approximately 40 to 50 years ago via bird dispersal of seeds that originated from Japan archipelago (Abe and Matsunaga, 2007; Sakai et al., 2010).
Vegetation analysis of the population showed that the tree layer was comprised of Neolitsea aciculata (Blume) Koidz. Dendropanax trifidus (Thunb.) Makino ex H. Hara and Ilex integra Thunb., and other species, and was dominated by Cinnamomum yabunikkei H. Ohba. In the subtree layer, S. praecox was the dominant species and Machilus thunbergii Siebold & Zucc. ex Meisn., Ligustrum japonicum, Camellia japonica L., Litsea japonica (Thunb.) Juss., Ficus erecta Thunb., Neolitsea sericea (Blume) Koidz., Prunus serrulata Lindl. f. spontanea (E. H. Wilson) Chin S. Chang, Neolitsea aciculata (Blume) Koidz., and Eurya japonica Thunb. were the major species. The shrub layer was composed of Litsea japonica (Thunb.) Juss., Camellia japonica L., Dendropanax trifidus (Thunb.) Makino ex H. Hara, Stauntonia hexaphylla (Thunb.) Decne., Trachelospermum asiaticum (Siebold & Zucc.) Nakai, Sinomenium acutum (Thunb.) Rehder & E. H. Wilson, Hedera rhombea (Miq.) Siebold & Zucc. ex Bean, M. thunbergii Siebold & Zucc. ex Meisn., Osmanthus insularis Koidz., Aucuba japonica Thun, N. sericea (Blume) Koidz. and Callicarpa japonica Thunb., including S. praecox. The major species in the herb layer were Farfugium japonicum (L.) Kitam., Cyrtomium falcatum (L.f.) C. Presl, Ophiopogon jaburan (Siebold) Lodd., and Arisaema thunbergii Blume.