Hanji, Korean traditional paper, has been an important cultural element in Korean history for more than 1,500 years (Jeong, 2015). The hanji has been made primarily from the bast fiber of Broussonetia species, especially ‘daknamu.’ The taxonomic nature of ‘daknamu’ has been confusing, but recent studies suggested the hybrid between B. papyrifera (L.) L’Her. ex Vent. and B. monoica Hance, B. × kazinoki (≡ B. monoica × B. papyrifera, B. × hanjiana) is it (Kim et al., 1992; Okamoto, 2006; Yun and Kim, 2009; Ohba and Akiyama, 2014; Chung et al., 2017). Daknamu has been recognized as a hybrid due to its intermediate morphology between the two parental species, B. papyrifera and B. monoica (Kim et al., 1992; Okamoto, 2006; Yun and Kim, 2009; Ohba and Akiyama, 2014; Chung et al., 2017). However, no one has applied molecular markers to test this, except a recent report of simple sequence repeat (SSR) marker test for hybrid origin of Daknamu (Rhu et al., 2019a). In Japan, B. × kazinoki is called ‘kõzo.’ In China, B. × kazinoki is highly valued not only for its fiber, but also as non-conventional forage, food additives, and traditional medicine, and its transcriptome (Xianjin et al., 2014) and chloroplast (cp) genome (Xu et al., 2018) has been studied.
One of the parental species, B. papyrifera, has also long been utilized in making paper as its name implies (“paper mulberry”) and barkcloth, tapa, especially in Oceania. Regarding the spread and utilization of B. papyrifera in Oceania, Chang et al. (2015) demonstrated that Pacific paper mulberry originated in southern Taiwan, providing the first ethnobotanical support for the “out of Taiwan” hypothesis of Austronesian expansion. Chang et al. (2015) analyzed cp ndhF-rpl32 intergenic spacer to trace the origin of paper mulberry widely introduced and cultivated in Oceania. In Korea, B. papyrifera is also natively distributed (Kim et al., 1992; Yun and Kim, 2009), though restricted to the coastal area and islands, or low elevation of inland (Rhu et al., 2019b). In case of Japan, B. papyrifera is non-native and assumed as introduced when paper-making technique was transferred from Korea (Okamoto, 2006; Mizumura et al., 2017; Chung et al., 2017). It is called as ‘kazino-ki’ in Japan.
The other parental species, B. monoica, has long been wrongly treated as B. kazinoki, until Akiyama et al. (2013) lecotypified Siebold’s collection and Ohba and Akiyama (2014) made taxonomic treatment later. In Korea, this species has long been wrongly recognized as daknamu, until Yun and Kim (2009) corrected it to Aegidaknamu, likewise ‘hime-kõzo’ in Japan.
In this paper, three molecular markers, cp ndhF-rpl32 IGS region, nuclear (nu) ribosomal internal transcribed spacer (rITS) and TOPO6 gene, were applied to test if B. × kazinoki is a hybrid of B. papyrifera and B. monoica. To avoid confusion, the species concept and treatment of Chung et al. (2017) were followed. Cp ndhF-rpl32 IGS region has been successfully presented the origin of oceanic B. papyrifera and nu markers, especially of single copy genes, have been useful in determining parental species in Ficus, Moraceae (Kusumi et al., 2012). TOPO6 gene has successfully demonstrated the hybrid and polyploid origin Hordeum (Brassac et al., 2012). Also, a sex marker developed by Penailillo et al. (2016) was tested.
Materials and Methods
Broussonetia specimens deposited in the herbaria of the National Institute of Biological Resources (KB) and the Daegu University (DGU) were carefully examined and identified according to the works of Kim et al. (1992), Yun and Kim (2009), and Chung et al. (2017). For nomenclature of Broussonetia species in Korea, concept of Chung et al. (2017) was followed. For molecular study, eleven B. × kazinoki, five B. monoica, and six B. papyrifera specimens were selected (Table 1).
The dried herbarium tissue of the selected 22 samples was powdered using Qiagen TissueLyser II (Qiagen, Hilden, Germany) and total genomic DNA was extracted using DNeasy Plant Mini Kit (Qiagen). Considering the maternal inheritance of plastid DNA and bi-parental inheritance of nu DNA of flowering plants, one plastid marker, ndhF-rpl32 IGS region and two nu markers, rITS and TOPO6 gene, were amplified and sequenced.
For the amplification of the cp ndhF-rpl32 IGS region, we applied ndhF – rpl32-3R and rpl32-3F – rpl32R primer combinations developed by Chang et al. (2015) and Shaw et al. (2014), which produced 1,288–1,400 bp long sequences. For nu rITS region, we applied AB101–AB102 primer combination designed by Douzery et al. (1999), which amplifies specifically vascular plants, avoiding contamination by fungi. As there are multiple copies with different-sized indels present with the nu rITS region of B. monoica and B. × kazinoki, gel-purified PCR products were cloned with TOPcloner TA kit (Enzynomics, Inc., Daejeon, Korea) and 2 to 12 colonies per sample were sequenced, after checking the sequence by direct sequencing of the purified PCR product of nu rITS region. Contrary to the B. monoica and B. × kazinoki, the sequences of nuclear rITS region of B. papyrifera were successfully obtained by directly sequencing the PCR product. For TOPO6 gene, a nu gene for subunit B of topoisomerase VI present as single-copy locus in most plant group, the region between exon 8 and exon 10 was amplified with TOPO6_8F2_1683 (5′-AGGCATTRATITGGTCIAARATGAG-3′) and TOPO6_10R2_2136 (5′-GTRATRACAGCCATYTGICGCAT-3′) primer combination, slightly modified from TOP6_8F_1683 and TOP6_10R_2146 of Blattner (2016), respectively. PCR amplification was done with 35 cycles of denaturation at 95°C for 30 s – annealing for 30 s – and extension at 72°C for 1 min, after the initial denaturation at 95°C for 2 min and final extension at 72°C for 7 min after the cycle. The annealing temperatures for the ndhF-rpl32 IGS, nu rITS, and TOPO6 gene were 53°C, 60°C, and 55°C, respectively. As directly sequenced B. × kazinoki TOPO6 gene sequences contained polymorphic sites (see Results) and the TOPO6 gene sequences of B. monoica is a little bit unclean after the T9–12 motif in the intron 9 (see Results), four B. × kazinoki and three B. monoica TOPO6 PCR products were cloned. DiaStar Taq DNA Polymerase (SolGent Co., Ltd., Daejeon, Korea) was used for the PCR amplification. All the sequencing was done by Macrogen Korea, Inc. (Daejeon, Korea). Sequences were cleaned and contiged with Sequencher 4.9 (Gene Codes, Ann Arbor, MI, USA) and aligned with AliView (ver. 1.25) (Larsson, 2014). Data matrices were imported into PAUP*4.0a (Swofford, 2002) and analyzed using neighbor-joining by applying Kimura’s 2-paramter model and maximum parsimony criteria. For the nu rITS sequence data, neighbor-joining analysis was only done, as the search for the optimal maximum parsimony tree was unfinished. Based on the sequence data of the cp ndhF-rpl32 IGS, haplotype networks were manually estimated. Broussonetia kaempferi sequences are included whenever possible, as an outgroup.
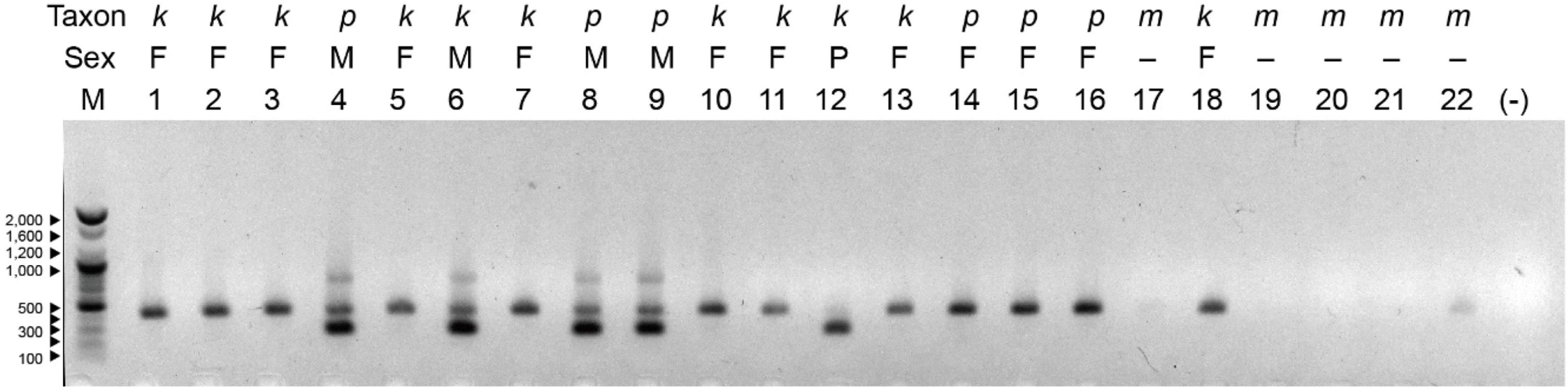
In addition, the utility of a sex-identity marker developed for B. papyrifera by Penailillo et al. (2016) was tested. Duplex PCR amplification using three primer combination, one forward and two reverse (MMRL and MMRS18) primers, was performed and the PCR products were electrophoresed on 1.5% agarose gel, together with 100 bp DNA ladder (Bioneer, Daejeon, Korea). Both male and female are supposed to produce 420 bp band (internal amplification control), while 273 bp band only for male individuals (male amplification).
Results
Morphological identification
Three Broussonetia species in Korea, B. papyrifera, B. monoica, and B. × kazinoki were clearly identified by the quantitative characters regarding the leaf, flower, and inflorescence type. All the samples are in flowering or bear fruits. As presented in Table 1, three female and three male samples were included for B. papyrifera. For B. monoica, four samples were monoecious, with both female and male inflorescences on the same individual, while one sample (Kim et al. Gageo25, VP288933), collected from Gageodo Isl., Sinan, Jeonnam is solely consisted of B. monoica type male inflorescences, assignable to B. kazinoki var. koreana sensu Yun and Kim (2009). For B. × kazinoki, ten female and one polygamous samples were included, as only female plants were available from KB and DGU. One specimen (Won 760), collected from Kurye, Jeonnam, was polygamous, with both female and male flowers in a short cylindrical inflorescence (Table 1).
Cp ndhF-rpl32 IGS region sequence analysis
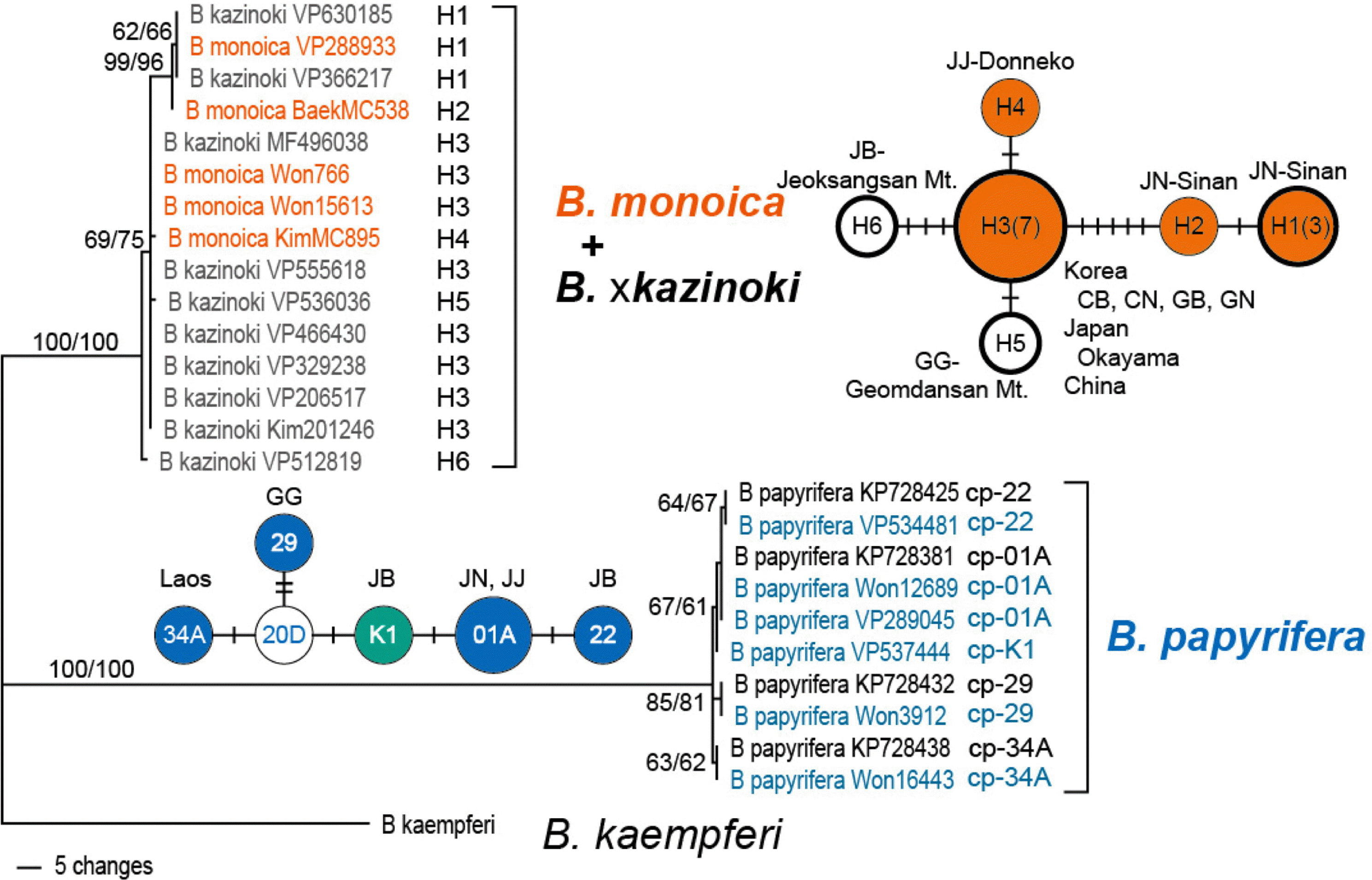
Cp ndhF-rpl32 IGS region of 20 samples were successfully amplified and sequenced, except for two samples, Won 2322 & Won 760 (Table 2). The length of the IGS region ranges from 1,220 to 1,337 bp; B. kaempferi is the longest as 1,337 bp, B. papyrifera 1,220–1,221 bp, and B. monoica and B. × kazinoki 1,320–1,322 bp long, respectively. The distinctively shorter length of B. papyrifera is caused by a 112 bp long indel and other 1–6 bp long indels, when compared to B. kaempferi. G + C content ranges 20.0–21.0%. When aligned, the data matrix was 1,467 bp long (Table 3). Phylogenetic analyses of the cp ndhF-rpl32 IGS region sequences clearly identified and separated the samples of B. papyrifera and B. kaempferi at species level, while samples of B. monoica and B. × kazinoki are clustered together (Fig. 1). When the newly obtained cp ndhF-rpl32 IGS region sequences of B. papyrifera were compared with those of Chang et al. (2015), five haplotypes, cp-01A, 22, 29, 34A, and K1, were recognized (Fig. 1, Table 2). Haplotype cp-K1 was recognized uniquely from a Korean sample for the first time, while the other four haplotypes were already reported from Chinese and Taiwanese samples by Chang et al. (2015). For B. monoica and B. × kazinoki, there were six haplotypes (H1–H6) recognized (Fig. 1, Table 2). Based on the sequence data, haplotype networks were manually estimated each for B. papyrifera, and B. monoica and B. × kazinoki haplotypes (Fig. 1). Haplotype cp-01A is detected from the two samples from Gageodo Isl., Sinan, Jeonnam and Jungmun Beach, Jeju of Korea and is distributed in Shanghai, Fujian, and Taiwan; haplotype cp-22 from Sinsido Isl., Gunsan, Jeonbuk and in Fujian, China; haplotype cp-29 from Daecheongdo Isl., Gyeonggi and in Guangdong and Fujian of China; haplotype cp-34A from Laos and in Yunnan, Guangxi, and Vietnam. The new haplotype cp-K1 was identified from the sample of Wido Isl., Buan-gun, Jeonbuk of Korea (Fig. 1, Table 2). In case of B. monoica and B. × kazinoki, haplotype H1 and H2 were detected from samples of Gageodo Isl. and Bigeumdo Isl. (H1), or Dochodo Isl. (H2) of Sinan, Jeonnam. Haplotype H3 is shared by both B. monoica and B. × kazinoki and shows most common and widest distribution from China, Japan, and diverse areas of Korea including Chungbuk, Chungnam, Gyungbuk, Gyungnam, Jeonbuk, and Jeonnam. Haplotype H4 from Seogwipo, Jeju, H5 from Mt. Geomdan, Hanam, Gyunggi, and H6 from Mt. Jeoksang, Muju, Jeonbuk (Fig. 1, Table 2). Four haplotypes, H1, H2, H3, and H4, were detected from B. monoica, while H1, H3, H5, and H6 from B. × kazinoki.
Nu rITS region sequence analysis
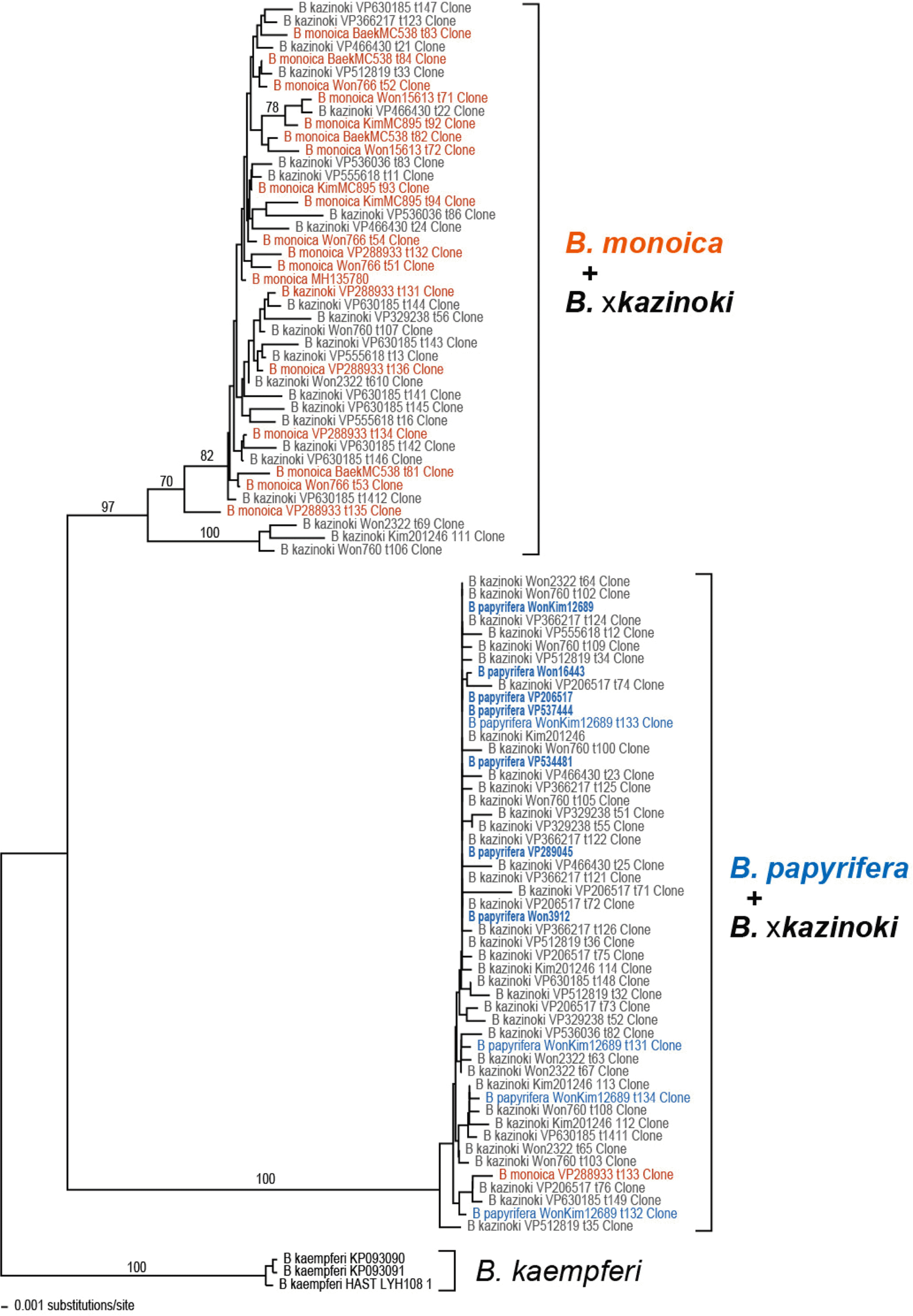
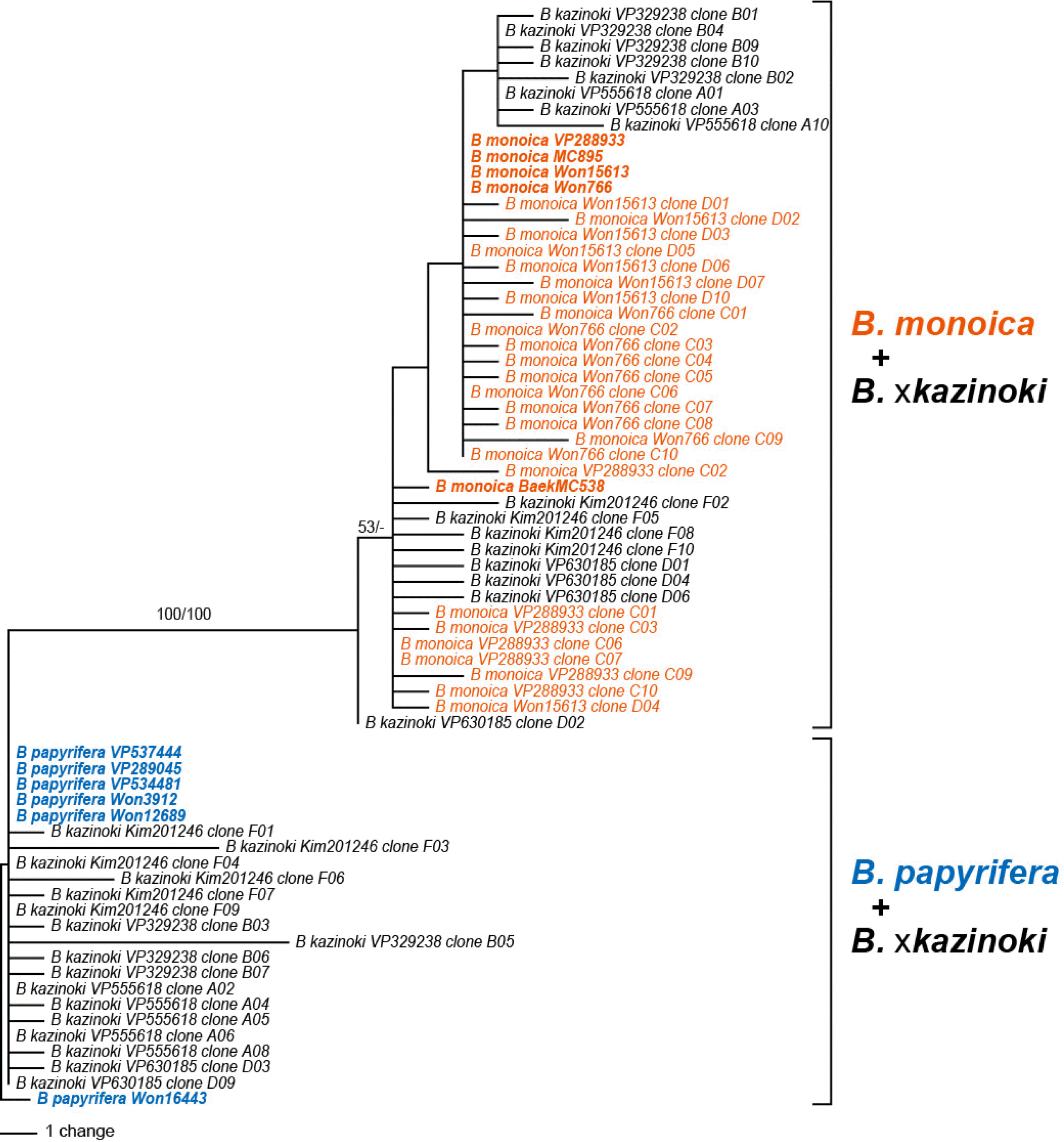
PCR amplification of the nu rITS region using the AB101–AB102 primer combination successfully amplified the marker. However, subsequent sequence determination by direct Sanger sequencing only worked for the samples of B. papyrifera, while the samples of B. monoica and B. × kazinoki required cloning due to the presence of multiple copies of nu rITS which caused unreadable sequences (Table 2). Therefore, seven directly sequenced sequences and 96 cloned sequences were obtained. Out of the 96 cloned sequences, 11 sequences were excluded from the further phylogenetic analysis because they were suspected as pseudogenic or chimeric sequences. In addition to the newly obtained sequence, one B. monoica sequence (MH135780) and three B. kaempferi sequences available from the NCBI GenBank were included for phylogenetic analysis. The length of nu ITS1 region ranges 225–247 bp, 5.8S rDNA 164–165 bp, and ITS2 region 236–262 bp, respectively. The G + C contents of nu ITS1 region ranges 68.0–71.8%, 5.8S rDNA 53.7–56.1%, and ITS2 region 67.2–72.4%, respectively. The aligned length of ITS1 + 5.8S + ITS2 region was 695 bp, while the data matrix was 962 bp long (Table 3). The cloned nu rITS sequences of B. × kazinoki are divided into two groups and clustered with B. papyrifera or B. monoica in the Neighbor-joining tree (Fig. 2). Except one sample, VP206517 where only six B. papyrifera type sequences were obtained, both the B. papyrifera and the B. monoica type sequences were obtained from the cloning of B. × kazinoki (Table 3). Also, the one B. monoica sample, Kim et al. Gageo25 (VP288933), had five B. monoica and one B. papyrifera type sequences.
Nu TOPO6 gene sequence analysis
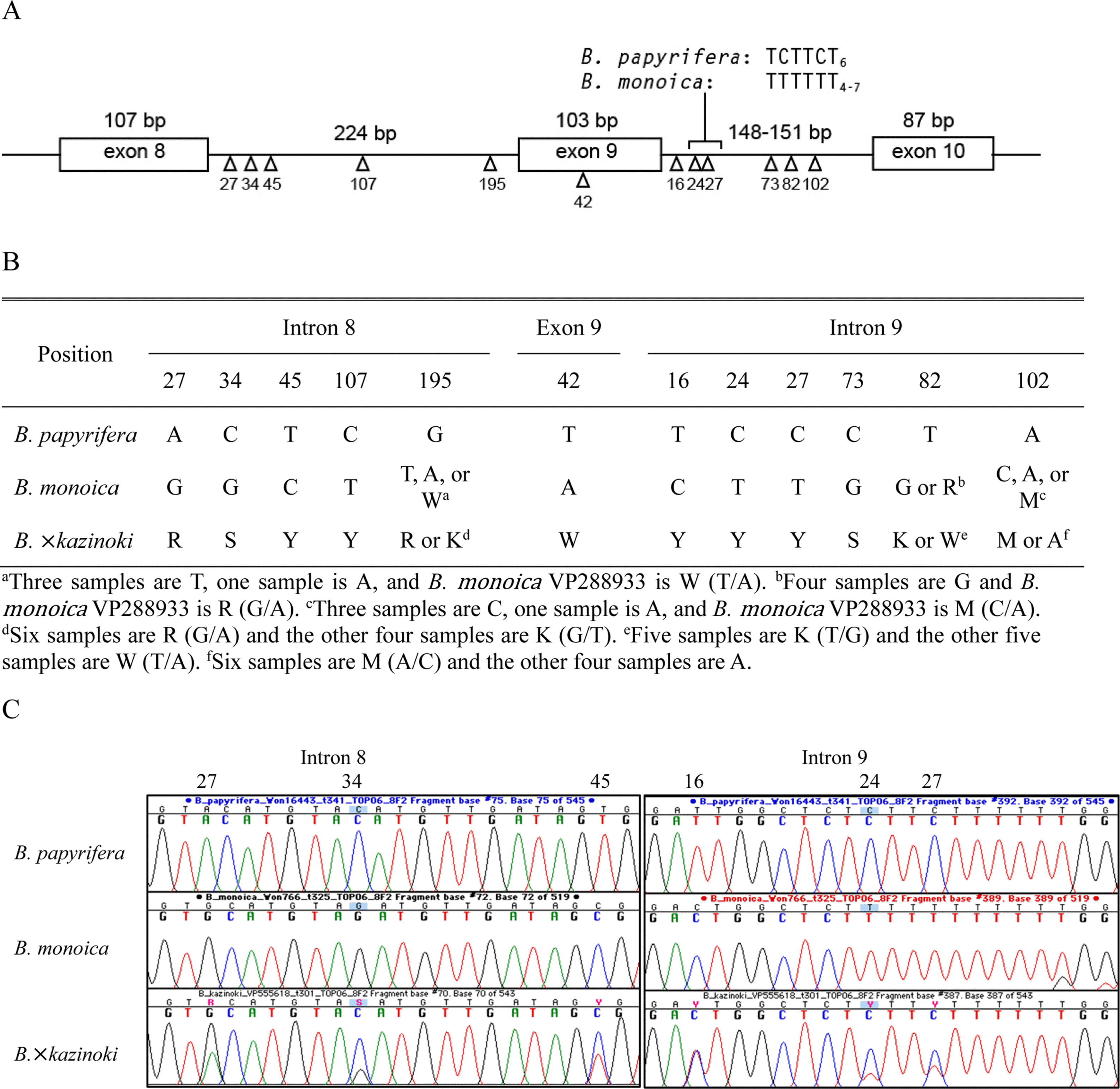
Region between exon 8 and exon 10 were successfully amplified and sequences were obtained by direct Sanger sequencing of the purified PCR products and subsequent cloning/sequencing of the PCR products (Figs. 3, 4, Table 2). The intron 8–exon 9–intron 9 region of nu TOPO6 gene is conserved in length for Broussonetia taxa included, except the T9–12 motif in the intron 9 (Fig. 3A). The G + C content ranged 35.1–37.0%. In case of B. monoica, and also for B. × kazinoki, the T9–12 mononucleotide repeat motif is present between the base positions 23 and 34 of the intron 9, while B. papyrifera has a TCTTCT6 motif (Fig. 3A). The T9–12 motif caused raised noise and ambiguities in sequence reading of B. monoica and B. × kazinoki immediately after it. There were five positions in intron 8, one position in exon 9, and six positions in intron 9, in total twelve nucleotide positions where the sequences of B. papyrifera and B. monoica are different (Fig. 3A, B). All the samples of B. ×kazinoki showed additivity in those 12 nucleotide positions where both the nucleotide peaks are overlapping (Fig. 3C). Subsequent cloning of B. × kazinoki TOPO6 gene PCR products were clearly separated into either B. papyrifera or B. monoica type sequences, while clones of B. monoica produced only B. monoica type sequences (Table 2). In case of B. monoica TOPO6 gene sequences, base position 195 of intron 8 (T or A), and base positions 82 (G or R) and 102 (C or A) of intron 9 show intraspecific sequence variation, while B. monoica VP288933 sample shows additivity in those bases (Fig. 3B).
The phylogenetic trees obtained from the maximum parsimony and the neighbor-joining are congruent in grouping the cloned sequences of B. × kazinoki TOPO6 gene into B. monoica or B. papyrifera clades (Fig. 4). The directly sequenced B. × kazinoki TOPO6 gene sequences and one putatively chimeric TOPO6 gene sequence were excluded from the phylogenetic analyses. Compared to the B. papyrifera + B. × kazinoki clade, sequences of B. monoica + B. × kazinoki clade present more sequence variation, probably due to the three intraspecific sequence variation.
Expression of sex marker
Sex marker was successfully amplified for B. papyrifera and B. × kazinoki, while none was amplified for B. monoica (Fig. 5, Table 2). Except for Won760 and Nam & Lee ND226 (VP206517), all the samples presented correct sex marker banding pattern (Fig. 5). Won 760 is a polygamous plant and produced only the male-specific band (273 bp), lacking the 420 bp internal control band. Nam & Lee ND226 (VP206517) has typical globose female inflorescences; however, it consistently produced male sex marker in two separate DNA extraction and PCR amplification repeats (Fig. 5, Table 2).
Discussion
Current study with molecular markers clearly proves that B. × kazinoki is a hybrid between B. monoica and B. papyrifera (≡ B. monoica × B. papyrifera), where maternal lineage is B. monoica and paternal is B. papyrifera, respectively. Cp marker ndhF-rpl32 IGS region demonstrated that the maternal lineage of B. × kazinoki shares haplotypes with B. monoica, while nu rITS and TOPO6 sequence types show that B. × kazinoki have both B. monoica and B. papyrifera copies. In addition to the molecular data, intermediate morphology of B. × kazinoki between B. monoica and B. papyrifera corroborate the hybrid origin of B. × kazinoki (Kim et al., 1992; Okamoto, 2006).
Although the sampling is limited, diversity and geographic distribution of haplotypes detected from B. × kazinoki and its maternal lineage B. monoica suggests that there may exist quite good amount of genetic diversity in natural populations of B. monoica, and therefore also in B. × kazinoki. Not all the four haplotypes detected from B. monoica are shared by B. × kazinoki, nor the four haplotypes detected from B. × kazinoki are shared by B. monoica. This suggests that there are more haplotypes in the wild than detected from this study, and there is good chance of finding diverse haplotypes when we increase sampling, as observed from the haplotype diversity of B. papyrifera (Chang et al., 2015). In the geographic distribution of haplotypes, the dominant haplotype H3 is shared by samples of Korea, Japan, and China, while haplotypes H1 and H2 are restricted to the southwestern islands of Korea, Dochodo Isl., Bigeumdo Isl., and Gageodo Isl. of Jeonnam, and H5 in Geomdansan Mt., Gyeonggi and H6 in Jeoksangsan Mt., Jeonbuk. The haplotype H3 may be a representative cultivar long been cultivated for paper-making in East Asia, while other B. × kazinoki haplotypes, H1, H5, and H6 are locally established and/or distributed. Further survey of cultivated B. × kazinoki haplotypes, together with B. monoica, in East Asia will improve our understanding of origin and usage history of B. × kazinoki in traditional paper-making.
The haplotypes of Korean B. papyrifera samples show that three out of four are shared with Chinese samples which are distributed in central and southern part of China such as Shanghai, Funjian, Guangdong, and Taiwan. Another haplotype K1 was solely detected from Sinsido Isl., Jeonbuk of Korea. Currently, B. papyrifera in Korea is mainly distributed along the coast and islands of Korea (Rhu et al., 2019b) and so does the sampling of the current study represent that: Korean B. papyrifera samples are from Jejudo Isl., Gageodo Isl., Wido Isl., Sinsido Isl., and Daecheongdo Isl. This shows that Korean B. papyrifera samples share the haplotypes commonly distributed in central and southeastern China, but not those of northern China, Japan, Indochina, and the Pacific (Chang et al., 2015). And the presence of Korean native type K1 suggests Korean B. papyrifera seems rather natural than introduced with paper-making. Therefore it is more reasonable to assume Korean B. × kazinoki is formed from the natural hybridization of the two parental species, rather than introduction from neighboring countries. Further sampling will be necessary to reveal the nature of Korean B. × kazinoki in cultivation whether they are selected from the wild or introduced.
According to Mizumura et al. (2017), interestingly, B. × kazinoki has been a major material for traditional paper in Japan for more than 1,200 years since its introduction to Japan by Dam Jing, a Korean Buddhist priest, in early 7th century. This suggests that the hybrid has long been available or introduced together with the technique in the earliest age of paper-making in Japan. As Ohwi (1965), Kitamura and Murata (1980), and Okamoto (2006) regarded B. papyrifera in Japan is non-native, B. × kazinoki in Japan utilized for paper-making may have been introduced elsewhere or formed later when B. papyrifera was introduced and cultivated in Japan. In case of Korea, B. papyrifera is regarded as native (Yun and Kim, 2009). Chang et al. (2015) reported three haplotypes from Japanese B. papyrifera, cp-05, 28, and 42, where cp-05 is restricted to Ryukyu Islands and Taiwan, cp-28 in Tokyo and Guangdong, China, and cp-42 only from Tokyo. Although the sampling of Japanese (Chang et al., 2015) and Korean B. papyrifera (in this study) is limited, the haplotypes detected show clear distinction in haplotypes and geographical distribution of them between the Korean and Japanese samples. Overall, current study supports the argument of Yun and Kim (2009) that B. papyrifera in Korea is native. On the contrary, B. papyrifera in Tokyo, Japan may have been introduced, while those in Ryukyu Islands are shared with eastern part of Taiwan. Still, further sampling of B. papyrifera is required to confirm the idea.
Broussonetia species are perennial and wind-pollinated, but also easily spread by root shoots asexually. When an interspecific hybrid is formed, the hybrid offspring easily become sterile. In case of B. × kazinoki, although no study on reproductive success of it has been done, the seed set and germination ratio is known to very low and therefore vegetative propagation methods such as root-cutting, cutting, and layering are recommended (http://iforest.nfcf.or.kr/; http://www.nongsaro.go.kr/portal/ps/psb/psbk/kidoContentsFileDownload.ps?kidofcomdtyNo=21395). However, although rarely, B. × kazinoki may be able to back-cross to the parental individuals and introgress as it can forms fruits and seeds. As the sex of B. × kazinoki is highly skewed to female over male (153 vs. 1 according to Rhu et al., 2019b), the female hybrid offspring will have more chance to introgress into the parental and hybrid individuals, while male, though rarely found, will also be able to play a role as pollen donor. Unfortunately, no study on the pollen viability of the male hybrid offspring has been done yet. The one sample, Kim et al. Gageo25 (288933), may have been formed by continued back-crossing of a male hybrid offspring with B. monoica, thereby showing six monoica-type and one papyrifera-type nu rITS sequence copies and combination of different TOPO6 sequence types of B. monoica.
The sex marker developed for B. papyrifera tested in this study showed that it works generally for B. papyrifera and B. × kazinoki, but there are two cases of escape for B. × kazinoki. For example, the polygamous individual of B. × kazinoki (Won 760) showed unusual banding pattern and Nam & Lee ND226 (VP206517), a typical female plants of B. × kazinoki morphologically, produced male sex marker. It is speculated that the sex marker region may have undergone recombination during the hybridization events, or the sex expression of B. × kazinoki is under control of other gene combinations. Therefore, it is suggested that application of sex marker for B. × kazinoki needs to be cautious.
Current study clearly demonstrated that B. × kazinoki is a hybrid between B. monoica and B. papyrifera, by applying cp and nu markers. Further sampling including neighboring countries and the study on reproduction may be required to fully understand the origin and history of B. × kazinoki utilized in paper-making.