Dalbergia L. f. (Fabaceae; Papilionoideae; Dalbergieae) is a pantropical tree genus with over 200 species that is distributed primarily in Central and South America, Africa, Madagascar, and Asia (Klitgaard and Lavin, 2005). Generally known as a rosewood (Klitgaard and Lavin, 2005; Patel, 2007; Vatanparast et al., 2013), its highly valuable timber products were historically imported into Korea as an important building material for furniture and statues in Buddhist temples during the Koryo Dynasty (Kim, 2013). However, we have recently found a population of Dalbergia hupeana Hance on the mountainous slopes of Mt. Yudal, Mokpo-si, Jeollanam-do, Korea (Figs. 1, 2).
Materials and Methods
The distribution of Dalbergia hupeana in Korea was investigated through field, literature, and oral surveys. Dried voucher specimens collected by the authors are being stored in the herbaria of Inha University (IUI) and the Chinese Academy of Sciences (PE), Beijing. Materials for morphological observations included dried specimens prepared from our own field samples at Mt. Yudal as well as those from the PE herbarium (Appendix). The molecular analysis involved DNA sequences of the nuclear internal transcribed spacer (nrITS) region plus plastid matK and rbcL (Table 1). Methods for analysis largely followed those described by Choi et al. (2013). Primer pairs were used for ITS (White et al., 1990), matK (Wojciechowski et al., 2004), and rbcL (Olmstead et al., 1992).
Results and Discussion
Surveys of habitat status and distribution
Plants of Dalbergia hupeana found at Mt. Yudal are located near the Buddhist temples. The population is restricted to lower elevations (approximately 120 m) where it dominates in more humid areas on slopes. This habitat contains more than 100 adult trees and numerous young seedlings of this species, with a maximum height of ca. 15 m and a diameter at breast height (DBH) of ca. 55 cm. Therefore, the D. hupeana population appears to be autochthonous and has a high rate of reproduction. The understory in this habitat also includes three evergreen species: Ligustrum japonicum Thunb., Trachelospermum asiaticum (Siebold & Zucc.) Nakai, and Hedera rhombea (Miq.) Siebold & Zucc. ex Bean. Further investigations revealed no other natural populations in Korea. However, trees of this species have been planted at three locations in Jeollado: Chonnam National University, Namwon National Gugak Center, and Manchui Garden. Among these, Manchui Garden is the oldest, having been built in 1913 at the end of the Chosun Dynasty. That site also features the oldest D. hupeana individual, a tree with a DBH of ca. 55 cm.
Morphological identification and characteristics
Based on descriptions of Korean floras, this species is most similar to the little known Korean Cladrastis platycarpa (Maxim.) Makino, which is considered indigenous to the Mt. Yudal region (Lee, 2003; Chang et al., 2011; Chang et al., 2012). However, the trees we found at Mt. Yudal clearly differ morphologically from C. platycarpa. The former is characterized by leaflets with obtuse or slightly emarginated apex, whitish corolla with purple stripes, and diadelphous (5+5) stamens, whereas the latter has leaflets with acuminate apex, whitish corolla with yellow spot in throat, and 10 free stamens. Alternatively, they show minor contrasts in characteristics when compared with descriptions of D. hupeana from China (Chen, 1994; Chen et al., 2010). In fact, our observations are in accord with the type specimen and protologue reported by Hance (1882). The Flora of China (Chen et al., 2010) states that both surfaces of D. hupeana leaflets are glabrous and have a sub-leathery texture. Moreover, their flowers are 6 to 7 mm long. However, leaves from plants collected at Mt. Yudal are pubescent on both sides, a papery or sub-leathery surface texture, and floral size of 8 to 10 mm. Although there is distinction between the description of the Flora of China and Mt. Yudal specimens, all the specimens we observed from the PE herbarium presented variable traits on the leaflet pubescence and texture. Hair and texture characteristics vary by season, with leaflets being pubescent on both sides and having a papery texture during the flowering season but being glabrescent on the adaxial surface and having a coriaceous texture during the fruiting period. We attribute those variations in flower sizes to different criteria being used for measurements. Therefore, we have devised an amended description for Korean D. hupeana.
Molecular characters from nrITS and cpDNA
The sequenced ITS region is 614 bp long and contains ITS 1 (231 bp), 5.8S (167 bp), and ITS 2 (216 bp). The ITS sequence shows low similarity (72.6% pairwise identity) to Cladrastis platycarpa (GenBank No. JQ676974). Blast results indicate high similarity to Dalbergia species. Both of those findings support our identification. Because complete, available sequences are not sufficient for D. hupeana and related species, we compared the partial ITS, matK, and rbcL regions and were able to distinguish D. hupeana from other congeneric species (Table 1). Only a single substitution occurred in the ITS 2 region while the matK and rbcL sequences were identical between Chinese and Korean D. hupeana.
The origin of Dalbergia hupeana in Korea: Native vs. Introduced
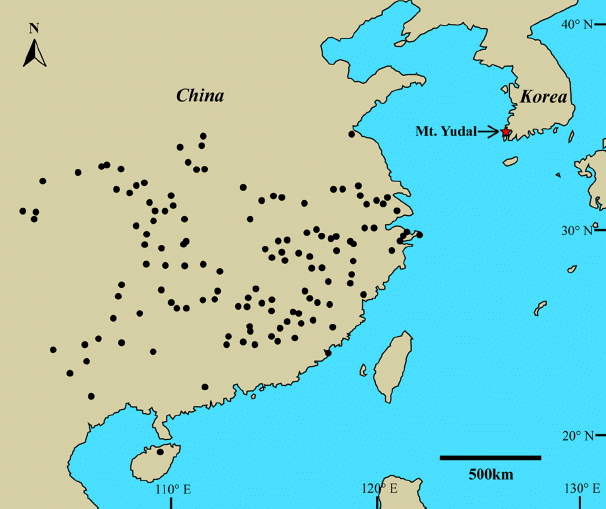
Researchers can speculate about whether D. hupeana is native or was introduced into Korea. However, the large, wide trunks of some trees suggest a persistent presence in Korea that might span at least one century. Plants of this species are naturally distributed throughout the Chinese provinces of Anhui, Fujian, Guangdong, Guangxi, Henan, Hubei, Hunan, Jiangsu, Jiangxi, Shandong, Shaanxi, Sichuan, Yunnan, and Zhejiang (Chen, 1994; Chen et al., 2010). Its northern boundary of range is thought to be approximately 34° 40′ N (Fig. 3). Therefore, a location at Mt. Yudal (34° 47′ N) would appear to be suitable as a natural habitat. The area near Mt. Yudal is also the only place in Korea that can sustain the growth of another Fabaceae family member, i.e., the warm-temperate tree Albizia kalkora Prain (Choi, 2007). From a biogeographic perspective, moreover, the pantropical distribution and mismatch between geographical and phylogenetic groupings of Dalbergia species may have resulted from long-distance dispersal via wind and water (Vatanparast et al., 2013). However, when we take into account its preferred environment, the distribution of this species in Korea appears to be more the outcome of a historically anthropogenic introduction. Our theory is also supported by the discovery of Dalbergia timber that dates from A.D. 1323 near the seabed at Mt. Yudal (Kim, 2013). Furthermore, neither morphological nor molecular data indicate that it is significantly different from Chinese plants, which implies that this disjunctive distribution is relatively recent. Whether native or introduced, it is true that these trees are now autochthonous and actively reproductive. Thus, populations of this warm-temperate species could potentially expand to more northerly locations in Korea due to global warming. Finally, the timing and manner of establishment for Korean populations could be further elucidated through the use of molecular phylogeographic analyses and extensive field surveys in the southwestern parts of Korea.
Shrubs, trees and climbing lianas. Leaves alternate, imparipinnate; leaflets alternate, rarely opposite, estipellate. Inflorescences terminal or axillary, racemes or panicles, usually numerous flowered. Flowers small; calyx campanulate, 5-toothed; teeth unequal, rarely subequal, lowest tooth usually largest, upper pair usually wider than others and partially connate; petals white, pale green, or rarely purple; standard ovate, oblong, or orbicular; wings with base cuneate, truncate, or sagittate; keel often boat-shaped, ± united above; stamens 9 or 10, monadelphous, usually united in an open sheath, or diadelphous and then in 2 bundles of 5 each (5+5), or 9 united and 1 free (9+1); anthers erect, small, dehiscent by short apical slits; ovary stipitate, few ovuled. Fruits oblong to oblongelliptical, samaroid, crescent or kidney-shaped, or orbicular to suborbicular, membranaceous, subcoriaceous or coriaceous, thickened over the seed cavity or thickened throughout. Seeds reniform, compressed; radicle inflexed.
Species: ca. 250
Distribution: pantropical; Africa (including Madagascar), Asia, Australia, South and Central America.
Dalbergia hupeana Hance, J. Bot. 20: 5. 1882; Chen, Fl. Reip. Popul. Sin. 40: 119. 1994; Chen et al., Fl. China. 10: 129. 2010. Type: Yichang, Hubei, China, T. Watters s.n. (herb. propr. n. 21189) (Holotype, BM, seen as a photo!)
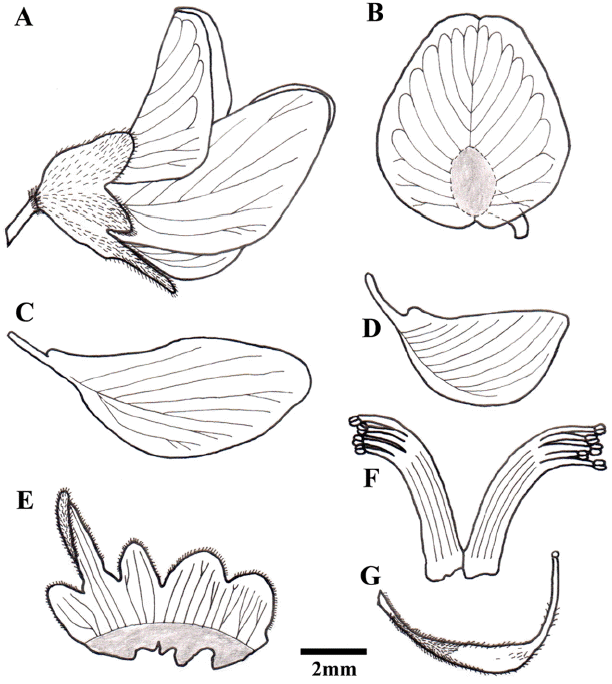
Tree, up to 15 m tall; bark dull gray or brownish gray; branches glabrous. Leaves of flowering twig 7- to 9-foliolate, 15–25 cm long including peduncle, sprouting in June; terminal leaflet elliptic to oblong elliptic, sometimes ovate or obovate, 5.0–7.5 × 2.5–3.5 cm, papery to subleathery, upper surfaces shiny, persistent sparse, short whitish hair, or occasionally glabrescent in autumn, lower surfaces pubescent with yellowish hair, veinlets prominent, base rounded or broadly cuneate, apex obtuse or slightly emarginated; lateral ones similar to terminal but gradually smaller toward peduncle. Panicles terminal or extending into axils of uppermost leaves, 5–20 × 5–20 cm, sparsely rusty, puberulent. Flowers 8–10 mm long; pedicel ca. 5 mm long, sparsely rusty, puberulent as calyx; bracts and bracteoles caducous, ovate, pubescent. Calyx campanulate, 5-lobed, 4.5–5.5 mm long including lowest lobe; two upper lobes rounded, subconnate, two lateral lobes smaller, ovate, lowest lobe lanceolate, ca. 2 times longer than others. Petals clawed; standard white with purple stripes inside, orbicular, emarginated at apex, 6.5–7.2 mm long, 5.3–6.3 mm wide; wings white, half-moon-shaped, 7.5–9.2 mm long, 3.2–3.8 mm wide; keel auriculate on upper side below, 6.0–6.7 mm long, 2.6–3.2 mm wide. Stamens 10, diadelphous (5+5). Ovary shortly stipitate, pubescent at base, 2- or 3-ovuled; style slender; stigma capitate, small. Fruit oblong or broadly ligulate, 4–7 (−12) × 1.3–1.5 (−1.8) cm, thinly leathery, reticulate opposite 1 or 2 (−4) seeds, base gradually tapering to stipe, apex acute. Seeds reniform, 7–14 × 5–9 mm. Fl. Jul–Aug. Fr. Sep–Oct.
Taxonomic note: This report of Dalbergia hupeana in Korea extends the distribution of this species northeastward. Moreover, this genus is a new addition to Korean flora. Its leaflet morphology, especially texture, is highly plastic by season, and lengths vary between flowering (5.0–7.5 cm) and non-flowering (7–11 cm) material. In Korea, new leaves emerge in late June, with the last dates being recorded for trees growing at Mt. Yudal. Based on this, the species can be readily distinguished from others because trees of the former appear to be withered during the springtime.
From the same locality at Mokpo-si, Lee (2003), Chang et al. (2011), and Chang et al. (2012) described Cladrastis platycarpa without including pictures or whole illustrations. However, leaf diagram for trees labeled as Korean C. platycarpa (Chang et al., 2011; Chang et al., 2012) did not resemble C. platycarpa but, rather, D. hupeana. Furthermore, we found no specimens of C. platycarpa during our field survey at Mt. Yudal. Therefore, the recording of C. platycarpa in Korean flora (Lee, 2003; Chang et al., 2011; Chang et al., 2012) is referable Korean D. hupeana.