나도국수나무족의 분류: 산국수나무속
Taxonomy of tribe Neillieae (Rosaceae): Physocarpus
Article information
Abstract
장미과 나도국수나무족(Neillieae)은 많은 분류학적 문제점을 안고 있는 나도국수나무속(Neillia), 국수나무속(Stephanandra), 산국수나무속(Physocarpus)의 3속에 속하는 약 18종으로 구성되어 있다. 나도국수나무족은 숙존하거나 조락하는 탁엽, 열편으로 갈리는 잎, 난형이고 광택이 있는 종자 및 풍부한 배유를 갖는 특징에 의해 장미과 내의 다른 족들과 뚜렷이 구분되며, 분자계통학적 연구 결과에서도 단계통군으로 강한 지지를 받는다. 그러나, 나도국수나무족 내의 종 간에 나타나는 복잡한 형태 변이 양상으로 인해 다양한 분류체계가 제시되어 있고 많은 종들이 기재되어 있으나, 전세계에 분포하는 나도국수나무족의 모든 종을 포함한 종합적인 분류학적 연구는 수행된 바 없다. 따라서 본 연구에서는 나도국수나무족의 종속지적 연구의 일환으로 기준표본을 포함한 표본 조사 및 야외 조사를 바탕으로 한 형태형질 분석 결과와 현재까지 밝혀진 분자계통학적 연구 결과를 토대로 산국수나무속의 분류를 제시하였다. 각 종에 대한 검색표, 모든 학명에 대한 자세한 명명법적인 검증 및 처리 그리고 나도국수나무족, 산국수나무속 및 각 종에 대한 기재, 분포도 및 조사한 표본의 목록을 제시하였다. 본 연구에서는 산국수나무속에 6종을 인식하였으며, 다음의 학명에 대한 선정기준표본을 지정하였다 (Opulaster pubescens, Opulaster ramaleyi, Spiraea opulifolia var. parvifolia, Spiraea opulifolia var. tomentella, Physocarpus michiganensis, Physocarpus missouriensis).
Trans Abstract
The tribe Neillieae, a small group of about 18 species in the Rosaceae, comprises three taxonomically difficult genera, Neillia, Physocarpus, and Stephanandra. The tribe, characterized by lobed leaves with persistent or deciduous stipules and ovoid, shiny seeds with copious endosperm, is strongly supported as a monophyletic group by a variety of lines of molecular evidence. Due to the high amount of morphological variation across the three genera and the species in tribe Neillieae, conflicting classification schemes and numerous species have been proposed over the past three centuries. However, no comprehensive systematic study of the group, including all species across their geographic ranges, has ever been undertaken. As part of a taxonomic revision of tribe Neillieae, a revision of Physocarpus based on the morphological examination of herbarium specimens, including types, and field observation is presented. Artificial keys, comprehensive nomenclatural treatments, descriptions, distribution maps, and lists of specimens examined are provided. Six species in Physocarpus are recognized. A lectotype is here designated for the following species: Opulaster pubescens, Opulaster ramaleyi, Spiraea opulifolia var. parvifolia, Spiraea opulifolia var. tomentella, Physocarpus michiganensis, and Physocarpus missouriensis.
The tribe Neillieae, a small group of about 18 species in the Rosaceae, has been treated as consisting of three taxonomically difficult genera, Neillia D. Don, Physocarpus (Cambess.) Raf., and Stephanandra Siebold & Zucc. (Maximowicz, 1879; Schulze-Menz, 1964). The tribe is characterized by lobed leaves with persistent or deciduous stipules, and ovoid shiny seeds with copious endosperm (Vidal, 1963). The monophyly of the tribe is strongly supported by chloroplast DNA sequence data, including rbcL (Morgan et al., 1994), matK and trnL genes (Potter et al., 2002), and combined six nuclear and four cpDNA data (Potter et al., 2007). The members of the tribe Neillieae are deciduous shrubs found mainly in mixed coniferous forests and riparian areas of the Himalayas, China, Korea, Japan and North America.
Many of the species of tribe Neillieae have horticultural merit. Their inflorescences and flowers are attractive and showy, and the plants are easily grown in gardens without special attention and can be propagated by cuttings. Because of these characteristics, several species such as N. sinensis Oliv., N. thibetica Bureau & Franch., P. capitatus (Pursh) Kuntze, P. opulifolius (L.) Maxim., and S. incisa (Thunb.) Zabel have been widely cultivated (Rehder, 1940; Cullen, 1971; Yu and Ku, 1974; Kruckeberg, 1982). Physocarpus opulifolius, native to eastern North America, has become naturalized in Europe and Asia (Ball, 1968; Lee, 1980).
The species of tribe Neillieae are native to eastern Asia and both western and eastern North America, exhibiting an intercontinental disjunct distribution pattern. This is one of several interesting biogeographic patterns that are found in the Northern Hemisphere (Raven, 1972; Wood, 1972), which have been the subject of taxonomic, biogeographic, and evolutionary studies over the past two centuries (Boufford and Spongberg, 1983; Manos and Donoghue, 2001; Manos and Meireles, 2015). The most plausible explanation for such a disjunct distribution pattern is that it is the product of more than one biogeographic event including multiple migrations between the two continents via the Bering and North Atlantic land bridges (Tiffney, 1985a, b; Manos and Donoghue, 2001; Manos and Meireles, 2015).
Oh and Potter (2005) assessed the historical biogeography of tribe Neillieae using a phylogenetic framework generated from multiple DNA sequence data. Their dispersal-vicariance analyses suggested that the most recent common ancestor of tribe Neillieae may have occupied eastern Asia and western North America and that the two major clades in tribe Neillieae (Physocarpus and Neillia/Stephanandra) may have been split by an intercontinental vicariance event. The species of Neillia and Stephanandra probably were diversified in eastern Asia, whereas the species of Physocarpus evolved in western North America (Oh and Potter, 2005). They also suggested that the eastern Asian species, P. amurensis (Maxim.) Maxim, was derived from an ancestor in western North America and that its current distribution resulted from dispersal across Beringia.
Tribe Neillieae has needed comprehensive systematic study. The morphological characters used to distinguish the three genera are variable, which has led taxonomists propose a number of conflicting classification schemes over the past three centuries. For instance, Bentham and Hooker (1865), Greene (1889), and Jones (1893) treated Physocarpus as part of the genus Neillia, with Stephanandra as a separate genus, whereas Kuntze (1891a) classified all species of tribe Neillieae in Physocarpus. In spite of the taxonomic confusion, the tribe as a whole has received little modern taxonomic attention. Vidal (1963) and Cullen (1971) published revisionary studies of Neillia and briefly discussed the morphological relationships among the three genera, but their studies concentrated mainly on the species of Neillia. Most treatments of Physocarpus and Stephanandra have been contributions to regional floristic manuals; consequently not all the species in each genus have been included (Rydberg, 1908; Fernald, 1970; Gleason and Cronquist, 1991; Yu and Ku, 1974; Holmgren, 1997).
Many species in tribe Neillieae are poorly known or exhibit considerable morphological variation. The patterns of variation within a species have not been examined comprehensively, leaving to the description of numerous segregate species. Species identities and relationships, as well as the correct application of taxonomic names have remained problematic (Robertson, 1974; Welsh, 1982).
Oh and Potter (2005) have investigated phylogenetic relationships among the three genera of tribe Neillieae using multiple DNA sequence data and have proposed that Neillia and Stephanandra should be merged into one genus. Their results indicate that the genus Physocarpus is a strongly supported monophyletic group and is sister to the Neillia/ Stephanandra clade. Their separate and combined analyses of sequence data from rDNA, cpDNA and LEAFY indicate that Stephanandra is nested within Neillia. The close relationship between Neillia and Stephanandra is also supported by morphological characteristics, such as their acuminate to caudate leaf apices, racemose or paniculate inflorescences, and the possession of a single carpel per flower with the exception of N. affinis Hemsl. var. polygyna Cardot ex J. E. Vidal, which has 3−5 carpels per flower. Stephanandra has been included in Neillia (Oh, 2006).
In a series of two papers, a comprehensive taxonomy of tribe Neillieae reflecting most recent morphological and molecular analysis of the group (Oh and Potter, 2003, 2005; Oh, 2004, 2006; Oh et al., 2010) is provided. As the first part of the revision, this paper provides taxonomic history of tribe Neillieae and taxonomic treatment of Physocarpus, including detailed nomenclature and morphological descriptions. The second part of the revision will cover Neillia.
Materials and Methods
More than 3,000 herbarium specimens were examined for this study. Specimens were borrowed from major herbaria, including A, BM, CS, E, GH, JEPS, K, KPM, L, MO, NEBC, NY, P, PE, POM, RM, UC, and UMO. Specimens at DAV, KB, and KUN, and those collected by the author were also examined. The data presented here, including plant measurements and distributions, were obtained mainly from herbarium specimens. Where more than 100 herbarium specimens were studied for a taxon, a subset of the total specimens examined is cited.
Nomenclatural History of Tribe Neillieae
The first published description of a species that can be assigned to the tribe Neillieae appeared before the time of Linnaeus in Tournefort (1700) under the genus Spiraea (S. opulifolia). Linnaeus, in his Species Plantarum (1753), included 11 species in Spiraea L. including S. opulifolia. After that, three additional species now assignable to tribe Neillieae were described in Spiraea: S. incisa Thunb. (1784), S. capitata Pursh (1814), and S. monogyna Torr. (1827).
Cambessèdes (1824) examined morphological variation among the species in the genus Spiraea that had been described by 1824, and recognized five sections in Spiraea (Spiraea, Ulmaria, Physocarpos, Gillenia, and Kerria). He placed S. opulifolia in the monotypic section Physocarpos, based on the presence of stipules and inflated follicles; however, the two other species now assigned to tribe Neillieae (S. capitata and S. incisa) were classified in section Spiraea. De Candolle (1825) also followed the Cambessèdes’ classification for those species, in which S. opulifolia was placed in section Physocarpos and both S. capitata and S. incisa were in section Spiraea.
Don (1825) first established the genus Neillia based on Wallich collections from Nepal and described two new species (N. thyrsiflora and N. rubiflora). Blume in the next year (1826) described a new monotypic genus Adenilema from Java in Indonesia, but he later synonymized it with Neillia (Blume 1855), as did Miquel (1855). Miquel, in his Flora van Nederlandsch Indië (1855), proposed a new name Neilliaceae, which included the genus Neillia only with N. thyrsiflora and N. rubiflora, while S. incisa was placed under the genus Spiraea. Meanwhile, Siebold and Zuccarini segregated a new genus Stephanandra in 1843 based on S. flexuosa, which has been treated as a taxonomic synonym of S. incisa (Thunb.) Zabel.
Medikus (1799) segregated Opulaster from Spiraea based on S. opulifolia, but this new name was invalid because neither a description nor a citation of previous work accompanied in the original publication. In 1832, Rafinesque published a new genus Epicostorus, based on Spiraea monogyna; however no subsequent authors ever used the name. Rafinesque (1838) also established Physocarpa by elevating Cambessèdes’ Spiraea section Physocarpos to generic rank. Rafinesque (1838) argued that Physocarpa was distinguihsed from Epicostorus by its bracts and swollen fruits (Epicostorus lacked bracts and had compressed, not swollen, fruits). The orthography of Physocarpa was modified to Physocarpus by Maximowicz (1879) and because of this, the authorship of the genus name has been confused.
Maximowicz (1879) was the first to group the three genera (Physocarpus, Neillia, and Stephanandra) in the tribe Neillieae. He distinguished the tribe Neillieae by its lobed leaves, stipules, and hard, shiny seeds with abundant endosperm. He also considered that each of the three genera was distinct. However, before this treatment, classifications of the three genera were not consistent. For example, Bentham and Hooker (1865) united Neillia and Physocarpus, while Stephanandra was treated as separate genus. Kuntze (1891a), on the other hand, placed all species of tribe Neillieae in the genus Physocarpus in his first part of Revisio generum plantarum. In the second part of the same book, he transferred all the species to the genus Opulaster, realizing that the name Opulaster was the earliest name for species of tribe Neillieae (Kuntze, 1891b).
The name Opulaster is listed in “nomina generica conservanda et rejicienda” as a rejected name against Physocarpus in the Vienna Code (Briquet, 1906). In spite of the rejection of the name Opulaster (Briquet, 1906), Opulaster was frequently adopted in floras and taxonomic treatments (Schneider, 1905; Rydberg, 1908, 1922; Small, 1933).
Rickett and Stafleu (1959) revised the list of “nomina generica conservanda et rejicienda” and noted that there was no need to conserve Physocarpus against Opulaster because Opulaster was a nomen nudum. They instead proposed an orthographical conservation in arguing that Physocarpus (Cambess.) Raf. should be conserved over Physocarpa Raf.
In 1972, Robertson proposed that the name Physocarpus (Cambess.) Raf. should be conserved against Epicostorus, to avoid unnecessary taxonomic confusion. Under the rules of the ICN (McNeill et al., 2012), if the two genera were united, Epicostorus would have to be retained because it is the earlier name. While Physocarpus has been widely used in the botanical and horticultural literatures, the name Epicostorus has never been used other than by the original author. This proposal was approved by the General Committee for Spermatophyta (McVaugh, 1973), and the conservation appeared in the Leningrad Code and onwards.
Most modern classifications of the tribe Neillieae have followed the circumscription of Maximowicz (1879), in which tribe Neillieae consists of Neillia, Physocarpus, and Stephanandra (Vidal, 1963; Schulze-Menz, 1964; Cullen, 1971; Kalkman, 1988; Takhtajan, 1997). Hutchinson (1964) added Guamatela Donn. Sm., a monotypic genus described based on specimens from Guatemala, in the tribe Neillieae. A recent phylogenetic study (Oh and Potter, 2006) using chloroplast DNA sequence data (rbcL, atpB, and matK) revealed that the genus is not a member of the Rosaceae, but belongs to Crossomatales, forming a distinct clade not nested in any of the families in the order. A new monotypic family Guamatelaceae was established to include Guamatela (Oh and Potter, 2006).
Taxonomic Treatment
Neillieae Maxim., Trudy Imp. S.-Peterburgsk. Bot. Sada 6: 216. 1879.−TYPE: Neillia D. Don
Unarmed deciduous shrubs. Leaves simple, alternate, the blade palmately veined, distinctly petiolate; stipules 2, opposite, free. Inflorescences corymbs, racemes, or panicles of racemes. Flowers bisexual; hypanthium cupulate, turbinate, campanulate, or cylindric; sepals 5, persistent; petals 5, deciduous; gynoecium apocarpous, carpels 1−5, free from hypanthium, the style terminal, the stigma capitate; ovules 2 or more per carpel. Fruit a follicle; seeds ovoidal, hardened, shiny, with copious endosperm. Base chromosome number: x = 9.
Neillieae was considered to be closely related to tribe Spiraeae (Kalkman, 1988). Phylogenetic analyses using DNA sequences (Morgan et al., 1994; Potter et al., 2002, 2007) do not support the close relationship of tribe Spiraeae to Neillieae. Phylogenetic relationship of tribe Neillieae is unresolved.
Key to the genera of tribe Neillieae
1. Leaves covered with stellate trichomes of various density; inflorescences corymbs; follicles dehiscent along both ventral and dorsal sutures ······························· Physocarpus
1. Leaves covered with simple trichomes of various density; inflorescences racemes or panicles of racemes; follicles dehiscent along ventral suture only ·································· ······································ Neillia (including Stephanandra)
Physocarpus (Cambess.) Raf., New Fl. 3: 73, 1838. nom. & orth. cons.; Spiraea section Physocarpus Cambess., Ann. Sci. Nat. (Paris) 1: 385. 1824 (‘Physocarpos’); Neillia section Physocarpus (Cambess.) Benth. & Hook. f., Gen. pl. 1: 612. 1865.−TYPE: Spiraea opulifolia L. [≡ Physocarpus opulifolius (L.) Maxim.].
Opulaster Medik. ex C. K. Schneid., Ill. Handb. Laubholzk. 1: 442. 1905.−TYPE: Opulaster bullatus Medik.
Shrubs with erect or spreading principal branches and short lateral branches; the bark in several layers peeling off in strips; pubescence usually present, the trichomes stellate. Leaves of the flowering branches ovate, reniform, orbicular, or oval, palmately 3−5-lobed, the margins doubly serrate or crenate; petioles evident; stipules linear to ovate, deciduous. Inflorescences terminating lateral branches of the season, bracteate, of umbel-like corymbs. Hypanthium shallowly cupulate, turbinate, or campanulate; sepals spreading when flowering, becoming erect after anthesis, ovate to triangular; petals spreading, orbicular, white or cream; stamens 20−40, exserted, arising from the rim of the hypanthium, some persistent; carpels 1−5, weakly connate at least at base, glabrous to moderately pubescent or densely pubescent, the style terminal; ovules 2−4 per carpel. Fruit a follicle, inflated or flattened, dehiscent along the ventral and dorsal sutures; seeds 1−4 per follicle, yellowish brown to light brown. Chromosome number: 2n = 18.
A recent phylogenetic analysis of Rosaceae (Oh et al., 2010) suggested that Physocarpus insularis (Nakai) Nakai, recognized as an endemic species on Ulleungdo Island in East Sea of Korea (Nakai, 1918; Lee, 1980; Lee, 2007), is not a member of the genus Physocarpus but that it should belong to the genus Spiraea. It has been formally transferred to Spiraea, S. insularis (Nakai) H. Shin, Y. D. Kim & S. H. Oh, based on morphology and DNA sequence data (Shin et al., 2011). Six species of Physocarpus are recognized in this treatment (Fig. 1).

Photographs of species of Physocarpus. A. P. alternans, B. P. amurensis, photo courtesy of Jin-Oh Hyun, C. P. capitatus, flowers, D. P. capitatus, fruits, E. P. malvaceus, F. P. monogynus, G. P. opulifolius, flowers, H. P. opulifolius, fruits. Note that P. capitatus and P. opulifolius have highly inflated fruits that are exposed from hypanthium (D and H).
Key to the species of Physocarpus
1. Carpels 1 or 2, rarely 3, follicles not highly inflated at maturity, more or less enclosed in the hypanthium and the sepals.
2. Corymbs with usually 1−8 flowers; carpels 1, rarely 2 ································································ 1. P. alternans
2. Corymbs with more than 10 flowers; carpels 2, rarely 3.
3. Follicles flattened laterally at maturity with more or less erect beaks, weakly connate above the middle of their length ·················· 4. P. malvaceus
3. Follicles not flattened at maturity with spreading beaks, weakly connate at the base.
4. Leaf blades longer than 4 cm; follicles 5.5−8 mm long; eastern Asia ····················· 2. P. amurensis
4. Leaf blades shorter than 4 cm; follicles 4–5.5 mm long; western U.S.A ·· 5. P. monogynus
1. Carpels 3−5, follicles highly inflated at maturity, more than twice as long as the hypanthium and the sepals.
5. Leaf blades of the flowering branches ovate, truncate to cordate at the base; teeth acute or acuminate; western North America ······················ 3. P. capitatus
5. Leaf blades of the flowering branches oval to ovate, attenuate to rounded, rarely truncate at the base; teeth more or less obtuse; eastern North America ············· ······························································ 6. P. opulifolius
1. Physocarpus alternans (M. E. Jones) J. T. Howell, Proc. Calif. Acad. Sci. 20: 130, 1931 (Fig. 1A); Neillia monogyna (Torr.) Greene var. alternans M. E. Jones, Zoe 4: 42, 1893; Opulaster alternans (M. E. Jones) A. Heller, Cat. N. Amer. Pl., ed. 2. 5, 1900.−Type: U.S.A. Nevada: Schell Creek Moutains, Duck Creek alt. 7,300 ft., 30 Jun 1893, Jones s.n. (holotype, POM!). Howell (1931) made a new combination Physocarpus alternans subsp. typicus J. T. Howell based on the type, when he newly described two other subspecies of Physocarpus alternans. However, the name is illegitimate because he ought to repeat the species epithet, alternans, to make an autonym.
Physocarpus alternans subsp. panamintensis J. T. Howell, Proc. Calif. Acad. Sci. 20: 132, 1931; Physocarpus alternans var. panamintensis (J. T. Howell) Jeps., Fl. Calif. 2: 164, 1936.−TYPE: U.S.A. California: about 150 yards north of saddle between Johnson and Surprise canyon, alt. 8,500 ft., Panamint Range, without date, Howell 3942 (holotype, CAS).
Physocarpus alternans subsp. annulatus J. T. Howell, Proc. Calif. Acad. Sci. 20: 133, 1931; Physocarpus alternans var. annulatus (J. T. Howell) Jeps., Fl. Calif. 2: 164, 1936.−TYPE: U.S.A. California: Wyman Creek, White Mts, alt. 8,500 ft., 2 Jul 1926, Duran 1682 (holotype, CAS; isotypes, UC-2 sheets!).
Shrub to 1.5 m high. Leaves of the flowering branches with blade 0.9−2 cm long, 1−2 cm wide, outline shape ovate to reniform, shallowly 3-lobed, the apex rounded, the base cordate, the margin doubly crenate, the upper and lower surfaces densely pubescent with stellate trichomes; petioles 0.3−0.9 cm long; stipules triangular to ovate, 1−2.5 mm long, entire, caducous. Inflorescences 1−8 (10)-flowered, densely pubescent with stellate trichomes; bracts lanceolate, 1.5−2.4 mm long; pedicels 0.3−1 cm long. Hypanthium campanulate or turbinate, 1.7−3 mm deep, 3.2−5 mm wide at the rim, densely pubescent with stellate trichomes; sepals ovate, 1.8−3.3 mm long, obtuse to rounded at the apex, densely pubescent with stellate trichomes on both surfaces; petals orbicular, white or pale pink; stamens 20−30, the longest filaments 1.3−2.5 mm long; carpels 1 rarely 2, if 2, weakly connate at the base, densely pubescent with stellate trichomes. Follicles 3−4.5 mm long, somewhat flattened laterally, with spreading beaks; seeds 1−3 per follicle, 1.6−2.1 mm long, yellowish brown to light brown.
Distribution. Western U.S.A., from eastern California, Nevada, Utah, to western Colorado and southwestern Wyoming, northward to Idaho (Fig. 2); pinyon-juniper woodland of desert mountains; limestone outcrops; 1,300−3,050 m.

The distribution of P. alternans, P. capitatus, and P. opulifolius.
Physocarpus alternans is morphologically distinct in the genus in that it has very short flowering branches with a cluster of very small leaves, fewer flowers per inflorescence, and very short pedicels compared to other species in the genus. In addition to the reduced habit of the species, the flowers of P. alternans usually have only one carpel. The rest of the species in Physocarpus have two or three to five carpels fused at least at base.
Phylogenetic analysis of tribe Neillieae (Oh and Potter, 2005) using nuclear ITS and chloroplast DNA sequences showed that P. alternans is the first diverging lineage in Physocarpus, sister to the remaining species in the genus (Fig. 3). Oh and Potter (2005) suggested that the gynoecium character may have supported the placement of P. alternans as the basal lineage in the genus if the single carpel is a synapomorphy for Neillieae and the 2−5-carpel condition evolved in Physocarpus. All of species in Neillia except for N. affinis var. polygyna have a single carple. It is also possible that the unicarpellate conditions in P. alternans and the genus Neillia evolved independently, if the most common ancestor of tribe Neillieae has a multicarpellate gynoecium.
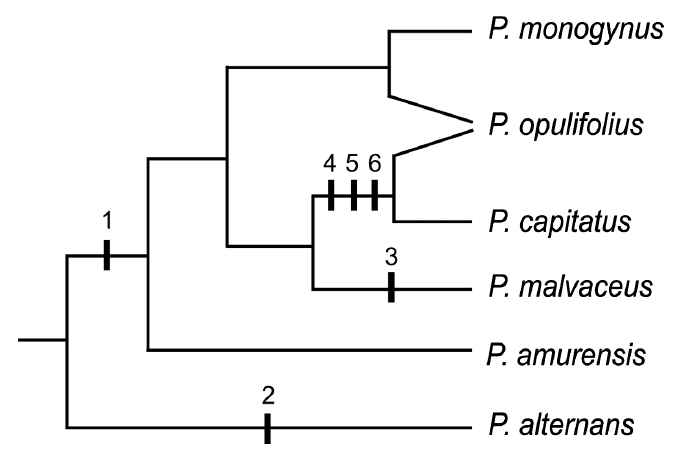
Phylogenetic summary tree of relationships in Physocarpus based on parsimony analyses of a combined chloroplast DNA (trnL-trnF, trnD-trnT, matK-trnK, and psbA-trnK), spacer regions of nrDNA (ITS and ETS), and the second intron of LEAFY. Apomorphies are indicated by number: 1 = 2-carpellate gynoecium; 2 = xerophytic habit; 3 = flattened follicle; 4 = 3−5-carpellate gynoecium; 5 = higly inflated follicle; and 6 = follicles exposed from hypanthium. Ancestral states of the characters for Physocarpus may be unicarpellate gynoecium, mesophytic condition, and slightly inflated follicle enclosed in hypanthium. Physocarpus opulifolius was suggested to be of hybrid origin bewteen P. capitatus and P. monogynus (Oh and Potter, 2005).
Representative specimens examined. U.S.A. California: Inyo Co., Tin Mtn, N side of summit, elev. 8,900 ft., 24 Jul 1978, DeDecker 4762 (UC); Mono Co., Indian Creek drainage, White Mts, elev. 8,750 ft., 29 Jul 1988, Morefield & Perala (NY). Colorado: Mesa Co., Colorado Nat. Mon., elev. 5,100 ft., sandstone cliff, 25 May 1943, Ripley & Barneby 5441 (NY); Moffat Co., Wild Mountain, ca. 0.3 mi SW of Burnt Springs, elev. 7,100 ft., 28 Jun 1988, Naumann 232 (CS). Idaho: Butte Co., The national reactor testing station, N of Howe, 5 mi to Black Canyon thence 2 canyons E, elev. 5,600 ft., 10 Jun 1967, Atwood 926 (NY); Twin Falls Co., Little Salmon river near Balanced Rock, 8 May 1936, Davis 18-36 (NY). Nevada: Clark Co., Grapevine Canyon, Sheep Range, elev. 6,400 ft., 8 Jun 1979, Ackerman 79-358 (NY); Elko Co., Pequop Mts, road leading S from I-80 to radio facility, W of Pequop summit, elev. 6,600 ft., 9 Jun 1981, Pinzl 4133a (GH); Esmeralda Co., Springdale Canyon on the E side, Lone Mt area, 14 mi W of Tonopah, elev. 8,800 ft., 30 Jul 1983, Tiehm & Wharff 8260 (NY); Lander Co., Toiyabe Range, head of Big & Kingston Creeks, 2 mi N of Bunker Hill, elev. 10,500 ft., 16 Jul 1981, Neese et al. 10752 (NY); Lincoln Co., Panaca Hills, at the uppper edge of Condor Canyon, elev. 5,500 ft., 23 Jun 1983, Tiehm & Williams 7967 (NY); Nye Co., Tybo Canyon, Hot Creek Range, 0.7 mi W of the site of Tybo, elev. 6,800 ft., 2 Jun 1987, Tiehm 11138 (NY); White Pine Co., Antelope Range, mouth of Cottonwood Canyon, 46 km airline distance E of Cherry Creek, elev. 2,010 m, 3 Jun 1984, Holmgren & Holmgren 10378 (NY). Utah: Box Elder Co., Regulator Canyon on the W side of Copper Mt, elev. 6,300 ft., 17 Sep 1984, Albee 5774 (NY); Cache Co., N side of Logan Canyon, elev. 5,800 ft., 20 May 1938, Maguire 16684 (UC); Carbon Co., Uinta Basin, on basin rim near Willow Springs, 10 mi E of Sunnyside, elev. 9,600 ft., 10 Jul 1935, Graham 9545 (GH); Daggett Co., ca. 7 mi SE of Manila, N flank of Boars Tusk Ridge, elev. 6,300 ft., 27 Jun 1979, Welsh & Moore 18702 (NY); Emery Co., 1 mi NW of Emery, elev. 7,700 ft., 12 Jun 1979, White 94 (NY); Garfield Co., Dixie National Forest, E upper slope of Sand Creek, elev. 7,800 ft., 29 May 1989, Franklin 6505 (NY); Grand Co., W of Dewey, elev. 4,200 ft., 5 May 1985, Franklin 1367 (NY); Juab Co., Granite Canyon, Deep Creek Mts, elev. 7,000 ft., 20 Jun 1933, Maguire & Becraft 2644 (GH, UC); Millard Co., Confusion Range, Skull Rock Pass, 1 Aug 1963, Welsh & Moore 2226 (NY-2 sheets); San Juan Co., Natural Bridges National Monument, White Canyon, ca. 1 mi E of Sipapu Bridge, 28 May 1968, Welsh & Moore 7149 (NY); Salt Lake Co., Salt Lake City, 22 Jul 1884, Leonard s.n. (NY); Uintah Co., Big Brush Creek Gorge, Uinta Mts, elev. 7,200 ft., 11 Jun 1993, Huber & Goodrich 62 (NY); Utah Co., Provo, Wasatch Mts, 16 Jun 1902, Goodding 1159 (A, NY, UC); Washington Co., Zion National Park, N side of Mesa top, Langston Mt, elev. 7,200 ft., 20 Jul 1988, Tuhy 3521 (NY). Wyoming: Sweetwater Co., Minnies Gap., elev. 6,400 ft., 4 Jul 1986, Dorn 4363 (NY).
2. Physocarpus amurensis (Maxim.) Maxim., Trudy Imp. S.-Peterburgsk. Bot. Sada 6: 221, 1879 (Fig. 1B); Spiraea amurensis Maxim., Mém. Acad. Imp. Sci. St.-Pétersbourg Divers Savants 9: 90, 1859; Opulaster amurensis (Maxim.) Kuntze, Revis. Gen. Pl. 2: 949, 1891.−TYPE: “Im Bureja-Gebirge, an felsigen nach Nord gelegenen Abhängen, eine Tagereise unterhalb Chinganskoi Piket, häufig,” 17 Aug 1856, Maximowicz s.n. (holotype, LE; possible isotype, MO!).
Physocarpus ribesifolius Kom., Izv. Bot. Sada Akad. Nauk SSSR 30: 202, 1932−Syntypes: Korea. “ad ripas fl. Jalu,” 4-16 Sep 1897, Komarov s.n. (LE); Russia. “Vladimir-Monomach in monte Taladza,” 30 May 1909, Djukina 64 (LE); “Vladimir-Monomach in monte Taladza,” 30 May 1909, Djukina 91 (LE); “in valle Kedrovaja padj, 7 Jul 1926, Popov s.n. (LE).
Shrub to 3 m high. Leaves of the flowering branches with blade 4−6.6 cm long, 3.5−5.8 cm wide, outline shape ovate to orbicular, shallowly 5-lobed, the apex acute, the base cordate, the margin doubly serrate, the teeth acute or acuminate, the upper surface glabrous, the lower surface glabrous to densely pubescent with stellate trichomes; petioles 1−2.5 cm long; stipules elliptic, 6 mm long, entire, caducous. Inflorescences more than 10-flowered, densely pubescent with stellate trichomes; bracts oblanceolate, 3−4 mm long; pedicels 1.5−1.8 cm long. Hypanthium campanulate, 2−4 mm deep, 5−5.5 mm wide at the rim, densely pubescent; sepals ovate, 3−4 mm long, acute at the apex, densely pubescent on both surfaces; petals orbicular, white; stamens 25−30, the longest filaments 4−5 mm long; carpels 2, rarely 3, weakly connate at the base, densely pubescent with stellate trichomes. Follicles 5.5−8 mm long, not inflated, with spreading beaks; seeds 2−3 per follicle, 1.5−1.8 mm long, yellowish brown to light brown.
Distribution. From northeastern China, North Korea to Primorskiy Kray and Amurskaya Oblast' in Russia, and to South Korea (Fig. 4); thickets in mixed coniferous forests, often rocky slopes of limestone; 1,400−2,000 m.

The distribution of P. amurensis.
This species is similar to P. monogynus and P. malvaceus in having two carpels connate at least at the base and follicles enclosed by hypanthium and sepals, but differs by having larger leaves with acute or acuminate teeth. Maximowicz (1859) stated that P. amurensis has four carpels. However, all specimens that I have seen, including a possible isotype of P. amurensis, have two, rarely three, carpels. In Physocarpus, when the follicles are mature, they dehisce along both ventral and dorsal sutures, generating two follicle walls with two beaks from one follicle. The type specimen contains only fruiting branches and the observation of four carpels made by Maximowicz (1859) may have resulted from erroneously counting the two follicle walls as two carpels.
I requested type specimens of P. amurensis and P. ribesifolius from LE, but did not receive any response. Poyarkova (1939) in “The Flora of the U.S.S.R.” stated that the type specimens for the two species are in LE. Poyarkova (1939) further mentioned that P. ribesifolia was described from the vicinity of Vladivostok, while Komarov (1932) did not specify the holotype for P. ribesifolius in the original description. I do not consider that Poyarkova’s statement is equivalent to designation of a lectotype for the species.
Moleular phylogenetic analysis (Oh and Potter, 2005) suggested that P. amurensis is sister to the North American species except for P. alternans (Fig. 3). The phylogenetic placements of P. alternans and P. amurensis, which has one and two to three carpels, repectively, as sucessive sisters to the remaining species of Physocarpus with 2 or 3−5 carpels. Biogegraphic analysis suggested that the Asian P. amurensis, nested within North American Physocarpus, might have been derived from western North American ancestor (Oh and Potter, 2005).
Representative specimens examined. CHINA. Hebei: eastern Tomb, Mt. Wuling-shan, 1,400-2,000 m, 10 Jun 1930, Li 11646 (A). Jilin: Helong, Sungseon, 456 m, 26 Sep 2013, Kim et al. C19-1 (KB).
KOREA. Hamgyeongbuk-do: in dumoisis Musan, 16 Jun 1914, Nakai 1791 (A); Yanggang-do: fluvium Jalu, Samsu circa Jang-su, 4 Jul 1897, Komarov s.n. (A). Kangwon-do: Gangneung-si, Mt. Seokbyung, 15 Sep 2011, Kim kjs130780 (KB–2 sheets).
RUSSIA. Primorskiy: Schkotovo Distr., “in fluxu superiore fl. Artemovka, in summitate monticuli Zmeinaja,” 15 Jun 1968, Gorovoi 6901 (NY); Chassan Distr., reservatum, Kedrovaja Padj, in summitate monticuli Czalban, 8 Jun 1975, Gorovoi 6902 (A, NY); Nakhodka enu, dry slopes and rocks of the “Brat” Mt., 24 Sep 1964, Gorovoi & Sakhno s.n. (UC); Partizansk Distr., "opp. Lozovoje, in declivibus petrosis montis Czandalaz,” 4 Aug 1980, Zhurba 6901a (A, NY). Yevreyskaya: Fluvium Kirma, Mons Londoko, 21 Aug 1895, Komarov 849 (NY).
3. Physocarpus capitatus (Pursh) Kuntze, Revis. Gen. Pl. 1: 219, 1891 (Fig. 1C and D); Spiraea capitata Pursh, Fl. Amer. Sept. 1: 342, 1814; Neillia capitata (Pursh) Greene, Pittonia 2: 28, 1889; Opulaster capitatus (Pursh) Kuntze, Revis. Gen. Pl. 2: 949, 1891; Opulaster opulifolius var. capitatus (Pursh) Jeps., Fl. W. Calif. 276, 1901.−TYPE: U.S.A. “Northwest coast of America,” Menzies s.n. (lectotype designated by Ewan, Editor’s Introduction for the facsimile of Fl. Amer. Sept. p. 106, 1979, BM!).
Spiraea opulifolia var. tomentella Ser. ex DC., Prodr. 2: 542, 1825; Opulaster opulifolius var. tomentellus (Ser. ex DC.) C. K. Schneid., Ill. Handb. Laubholzk. 1: 442, 1905; Physocarpus opulifolius var. tomentellus (Ser. ex DC.) B. Boivin, Naturaliste Canad. 93: 434, 1966.−TYPE: without specific locality and collector’s name (lectotype designated here, G-DC (IDC microfiches!); isolectotypes, G-DC (IDC microfiches–2 sheets!)). There are three original specimens for the taxon in G-DC. One labeled “Spiraea opulifolia tomentella Ser.” is selected here as the lectotype.
Spiraea opulifolia var. mollis Torr. & A. Gray, Fl. N. Amer. 1: 414, 1840; Neillia opulifolia var. mollis (Torr. & A. Gray) W. H. Brewer & S. Watson, Bot. California 1: 171, 1876.−TYPE: U.S.A. “Columbia,” Nuttall s.n. (holotype, NY!).
Spiraea ribifolia Nutt. ex Torr. & A. Gray, Fl. N. Amer. 1: 414, 1840.−TYPE: U.S.A. Oregon: Nuttall s.n. (holotype, NY!).
Spiraea opulifolia var. multiflora Durand, J. Acad. Nat. Sci. Philadelphia 3: 87, 1855.−TYPE: U.S.A. California: Nevada [city], Pratten s.n. (holotype, P!)
Opulaster cordatus Rydb., N. Amer. fl. 22: 242, 1908.−TYPE: U.S.A. Idaho: in a deep canyon, Farmington Landing, Lake Coeur d’Alene, 7 Jul 1892, Sandberg et al. 575 (holotype, NY!; isotypes, GH!, US).
Shrub to 2.5 m high. Leaves of the flowering branches with blade 3.2−8.5 cm long, 3.1−8.3 cm wide, outline shape ovate, shallowly 3-lobed, the apex acute, the base cordate, the margin doubly serrate, the teeth acute or acuminate, the upper and lower surfaces glabrous to moderately pubescent with stellate trichomes; petioles 1.1−3 cm long; stipules lanceolate, 5 mm long, entire, caducous. Inflorescences more than 10-flowered, sparsely to densely pubescent with stellate trichomes; bracts spatulate, 3−4 mm long; pedicels 0.9−2.6 cm long. Hypanthium cupulate to turbinate, 1.5−2.5 mm deep, 4−4.8 mm wide at the rim, densely pubescent with stellate trichomes; sepals ovate, 2.5−3.5 mm long, acute at the apex, densely pubescent with stellate trichomes on both surfaces; petals orbicular, white; stamens 25−30, the longest filaments 4−5 mm long; carpels 3−5, weakly connate at the base, glabrous to moderately pubescent. Follicles 8−10 mm long, highly inflated, with spreading beaks; seeds 2−3 per follicle, 2−2.5 mm long, yellowish brown to light brown. Chromosome number: n = 9 (Sax, 1931).
Distribution. Western North America, mainly coastal ranges and cismontane California, north through Oregon, Washington, British Columbia, and southern Alaska, disjunctively in northern Idaho (Fig. 2); mixed coniferous forests; along streams and river banks; 70−1,200 m.
Physocarpus capitatus is similar to P. opulifolius in having 3−5 carpels and highly inflated follicles which are more than twice as long as the hypanthium and the sepals at maturity. It is distinguishable from the latter in having leaves of the flowering branches that are ovate with truncate to cordate bases and marginal teeth that are acute or acuminate. The leaves of non-flowering branches in P. opulifolius are not significantly different from those in P. capitatus; thus they are not useful in distinguishing the two species. The distributions of the two species do not overlap. Physocarpus capitatus is confined to western North America, while P. opulifolius is widely distributed in eastern North America.
In his original description, Pursh mentioned another specimen collected by Lewis and Clark and indicated that he had observed the specimen in the Lewisian Herbarium. However, Reveal et al. (1999) examined the Lewis and Clark collections of vascular plants and discovered that the specimen mentioned by Pursh was not in PH. They suggested that it might be mixed with a Menzies collection. I requested the Lewis and Clark specimen from BM along with the Menzies specimen, but the Lewis and Clark collection was not located. The specimen collected by Menzies consisted of three branches. Pursh mentioned that the Lewis and Clark specimen was imperfect, but all three branches of the Menzies specimens bear leaves and inflorescences and are at the same developmental stage, suggesting that the Lewis and Clark collection is not part of the Menzies specimen. It appears that the Lewis and Clark collection is missing.
Holmgren (1997) placed Opulaster cordatus, which was described from Idaho, in synonymy with P. malvaceus. The type specimen of this name has flowers with five carpels that are connate only at the base and have moderately dense stellate pubescence. This places the type in P. capitatus.
Representative specimens examined. CANADA. British Columbia: Vancouver Island, Victoria, Sep 1903, Anderson s.n. (A); Goliath Bay, Jervis Islet, 11 Jul 1948, Bong 60 (UC–2 sheets); 14 mi ENE of Bella Coola, 6 Jul 1956, Calder et al. 18408 (UC); S end of small lake, ca. 1 mi N of Odlum, 18 Jun 1941, Hitchcock & Martin 7383 (A, NY, UC); Chilliwack Valley, 19 Jun 1901, Macoun 34364 (GH, NY); Canoona Lake, Princess Royal Island, 21 Jun 1936, McCabe 3536 (UC); Selkirk, near mouth of Downie Creek, 9 Aug 1905, Shaw 1115 (GH, NY–2 sheets).
U.S.A. Alaska: Prince of Wales Island, Salmon Bay Lake, elev. 100 ft., 22 Jun 1979, Perkins 268 (NY). California: Alameda Co., Berkeley, 4 Jun 1888, Greene s.n. (JEPS, NY); Amador Co., Jackson, elev. 1,500 ft., 30 Apr 1893, Hansen 236 (NY, UC); Butte Co., Little Chico Creek, May 1896, Austin 730 (UC); Contra Costa Co., near Lafayette, 13 Apr 1895, Davy 1000 (UC); Del Notre Co., Darlingtonia, Smith River, elev. 400 ft., Jun 1937, Parks & Parks 24064 (A, GH, NY, UC–2 sheets); El Dorado Co., 0.25 mi NW of Riverton, elev. 3,400 ft., 7 Jun 1934, Johannsen & Gifford 252 (UC); Humboldt Co., Mad River at S end of Mad River Bridge, 3 Jun 1977, Nelson & Baker 3387 (NY); Lake Co., Banks of Kelsey Creek, 6 mi S of Kelseyville, 26 Jun 1931, Schulthess s.n. (UC); Marin Co., 4.2 mi below Stinson Beach on Stinson Beach Road, 31 Jul 1941, McVaugh 6191 (A); Mariposa Co., Wawona, 11 Jul 1936, Mason 11205 (UC); Mendocino Co., Big River, Jun 1903, McMurphy 268 (NY); Napa Co., White Sulphur Spring, St. Helena, 12 May 1907, Chandler 7565 (UC); Plumas Co., along Hwy 70, 6 mi S from junction of Hwy 89 & 70, 5 Jun 1973, Kenney et al. 431 (NY–2 sheets); San Luis Obispo Co., upper Las Tablas Creek, 20 May 1969, Hoover 11318 (UC); San Mateo Co., foothills near Redwood City, 4 May 1902, Abrams 2397 (NY, UC); Shasta Co., Bailey Cove Trail, on the W side of the McCloud River Arm of Shasta Lake, elev. 1,100 ft., 27 Apr 1994, Oswald & Ahart 6140 (JEPS); Siskiyou Co., Wagon Creek on E side of Mt. Eddy, 14 Jul 1990, Ertter 9334 (NY, UC); Sonoma Co., Santa Rosa, 4 Jun 1902, Heller 5647 (GH, NY–2 sheets, UC); Tulare Co., Dorst Creek, Sequoia National Park, 10 Sep 1912, Fry 4 (JEPS); Tuolumne Co., Italian Bar, 16 May 1916, Grant 771 (JEPS); Trinity Co., Beside Dan Rice Creek and Scott Mtn Road, 3 mi S of Scott Mtn Forest Camp, elev. 4,550 ft., 3 Aug 1955, Barbe 341 (UC). Idaho: Benewah Co., S shore of Chatcolet Lake, along state Hwy 5, elev. 2,125 ft., 22 Jul 1985, Brunsfeld 2564 (NY); Idaho Co., S Fork Clearwater River, 20 Jun 1941, Davis 3368 (A, UC); Latah Co., 1 mi NE of bridge over Potlatch River, 2 air mi SE of Helmer, elev. 2,900 ft., 5 Jul 1986, Johnson 1645 (NY); Shoshone Co., Kit Price campgroud, along Coeur d’Alene River, alt, 2,650 ft., 22 Jun 1986, Johnson & Johnson 1611 (NY). Oregon: Benton Co., W of Corvallis, on Oak Geek Road, 29 Jun 1960, Dennis & Mason s.n. (GH, NY); Clackamas Co., Eagle Creek, 20 May 1928, Thompson 4268 (GH); Clatsop Co., saddle area of Saddle Mtn, 9 Jul 1962, French 2147 (NY); Crook Co., Eagle Creek, 27 Aug 1897, Coville & Applegate 693 (A); Curry Co., 4 mi above mouth of Rogue River, 17 Jun 1935, Keck & Clausen 3411 (UC); Douglas Co., bank of Calapooya River, elev. 800 ft., 25 Jul 1899, Barber 93 (GH); Hood River Co., 0.5 mi N of Lost Lake in Mt. Hood, 28 Aug 1927, Keck 318 (UC); Jackson Co., Ashland Creek, 1 mi S of Ashland, 8 Aug 1927, Mason 4045 (UC); Jefferson Co., Lake Suttle, elev. 4,000 ft., 23 Jun 1939, Hitchcock & Martin 4894a (A, NY, UC); Josephine Co., Siskiyou National Forest, 24 Jun 1973, Denton 2667 (NY); Lane Co., Junction of the Mckenzie and the Willamette River, N of Eugene, 29 May 1924, Constance s.n. (UC); Lincoln Co., Suislaw National Forest, along Tum Tum Road, elev. 350 m, 15 Jun 1995, Walker & Annable 1462 (NY); Marion Co., 4 mi N of Sublimity, 9 Aug 1941, McVaugh 6315 (A, UC); Multnomah Co., Portland, 11 Jun 1886, Drake & Dickson s.n. (NY); Yamhill Co., Walker flat, 11.2 mi air distance W of Carlton, elev. 552 m, 25 Jun 1985, Halse 3094 (UC); Washington Co., Hillsboro, 23 Aug 1911, Smith 4106 (NY); Wasco Co., Des Chutes Canyon near Maupin, 28 May 1933, Peck 17352 (NY). Washington: Clallam Co., Olympic Peninsula, along road to Sol-Duc, near Lake Crescent, 20 Jul 1960, Yuncker & Welch 18880 (NY, UC); Clark Co., Manor, 14 Jul 1899, Piper 2074 (NY); Grays Harbor Co., 5 mi SE of Humptulips, 13 Aug 1941, McVaugh 6363 (UC); Island Co., Whidbey Island, 12 Jul 1899, Garder s.n. (UC); King Co., Seattle, 18 Jul 1880, Engelmann & Sargent s.n. (A); Kitsap Co., Manchester, 17 Jun 1923, Warren 111 (NY); Kittitas Co., near Cle Elum, 14 Jul 1930, Palmer 37898 (A, NY); Lewis Co., moist meadows at western base of Storm King, elev. 3,000 ft., 28 Jun 1932, Thompson 8600 (A, GH, NY); Pierce Co., along Muck Creek, near Spanaway, 2 Jun 1961, Kruckeberg 5206 (UC); San Juan Co., Blakely Island, San Juan Islands, 25 Jun 1 - Aug 1917, Zeller & Zeller 1145 (A–2 sheets, GH, NY); Skagit Co., Skagit Delta, 14 Jul 1927, Mason 3952 (UC); Snohomish Co., Marysville, Jun 1927, Grant s.n. (A); Thurston Co., Millersylvania State Park, 10 mi S of Olympia, 15 Aug 1975, Taylor & Taylor 20108 (NY); Whatcom Co., Bellingham, 6 Jun 1916, Eugberg s.n. (NY); Yakima Co., Yakima region, in 1882, Brandegee s.n. (UC).
4. Physocarpus malvaceus (Greene) Kuntze, Revis. Gen. Pl. 1: 219, 1891 (Fig. 1E); Neillia malvacea Greene, Pittonia 2: 30, 1889; Opulaster malvaceus (Greene) Kuntze, Revis. Gen. Pl. 2: 949, 1893; Neillia monogyna (Torr.) Greene var. malvacea (Greene) M. E. Jones, Zoe 4: 43, 1893.−TYPE: U.S.A. Idaho: dry rocky banks above the northern shore of Lake Pend d’Oreille, 8 Aug 1889, Greene s.n. (holotype, NDG; isotypes, NY!, US).
Spiraea opulifolia var. pauciflora Nutt. ex Torr. & A. Gray, Fl. N. Amer. 1: 414, 1840, p.p.; Opulaster pauciflorus (Nutt. ex Torr. & A. Gray) A. Heller, Bull. Torrrey Bot. Club 25: 626, 1898; Physocarpus pauciflorus (Nutt. ex Torr. & A. Gray) Piper, Fl. Palouse Reg. 94, 1901.−TYPE: U.S.A. Oregon: Blue Mts., Nuttal s.n. (lectotype, NY!, designated by Heller, Bull. Torrey Bot. Club 25: 581, 1898). Torrey and Gray (1840) cited two specimens, one of which is the holotype of Spiraea monogyna (James s.n.). Heller (1898) designated the Nuttall collection from the Blue Mountains of Oregon as the lectotype.
Opulaster pubescens Rydb., Mem. New York Bot. Gard. 1: 205, 1900; Physocarpus pubescens (Rydb.) A. Nelson in J. M. Coulter and A. Nelson, Man. Bot. Rocky Mt. 248, 1909.−TYPE: U.S.A. Utah: Ogden, 8 May 1871, Porter s.n. (lectotype designated here: NY!). Syntypes: U.S.A. Montana: near Hound Creek, Rocky mountain sides, alt. 5,600 ft., 31 Jul 1883, Scribner 35 (NY!, GH?).
Shrub to 2 m high. Leaves of the flowering branches with blade 2.9−6.2 cm long, 2.6−5.9 cm wide, outline shape ovate to orbicular, shallowly 3-lobed, the apex acute, the base cordate, the margin doubly crenate, the upper surface glabrous, the lower surface sparsely pubescent with stellate trichomes; petioles 0.6−1.8 cm long; stipules elliptic, 4.5−7 mm long, entire, deciduous after anthesis. Inflorescences more than 10-flowered, densely pubescent; bracts spatulate, 3−4 mm long; pedicels 1.1−1.5 cm long. Hypanthium campanulate, 2.3−3 mm deep, 3.5−4 mm wide at the rim, densely pubescent with stellate trichomes; sepals ovate, 3−3.5 mm long, acute to obtuse at the apex, densely pubescent with stellate trichomes on both surfaces; petals orbicular, white or cream; stamens 25−30, the longest filaments 4−4.5 mm long; carpels 2, rarely 3, weakly connate to above the middle, densely pubescent. Follicles 6−7 mm long, flattened laterally, with more or less erect beaks; seeds 1−2 per follicle, 2−2.5 mm long, yellowish brown to light brown.
Distribution. Western North America, from southeastern British Columbia and southern Alberta through eastern Washington and Oregon, Idaho, western Montana and Wyoming, southward to northeastern Nevada and Utah (Fig. 5); mixed coniferous forests, Douglas fir, aspen, and sprucefir forests; rocky slopes, humid canyon slopes, and lake shores; locally predominant shrubs under coniferous forests in many localities; 500−2,900 m.
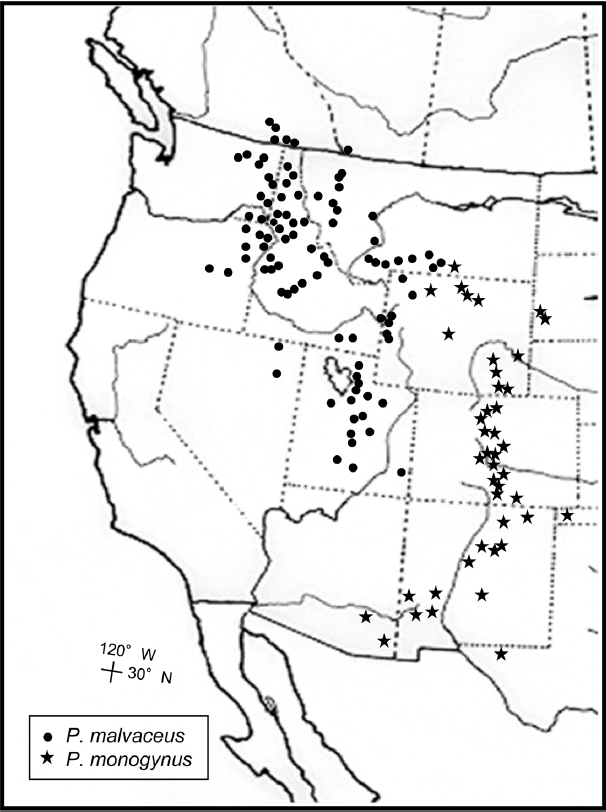
The distribution of P. malvaceus and P. monogynus.
Physocarpus malvaceus can be distinguished from other species of Physocarpus by having flattened follicles with more or less erect beaks at maturity. Greene (1889) mentioned that the fruits of this species are indehiscent and are similar to the silicle of a large Lepidium. However, the follicles dehisce along both ventral and dorsal sutures as in all other species of Physocarpus. As Jones (1893) noticed, they may be indehiscent in dry weather.
Because both P. malvaceus and P. monogynus have two carpels united at least at the base and both are distributed in the Rocky Mountain region, it is difficult to distinguish the two species at flowering stage. The two species, however, can be easily distinguished by fruit characters: P. malvaceus has laterally flattened follicles with more or less erect beaks at maturity, whereas P. monogynus has non-flattened follicles with more or less spreading beaks. Leaves of P. monogynus tend to more deeply lobed and smaller than those of P. malvaceus, although there is some variation within each species. Inflorescences of P. malvaceus are always densely stellate-pubescent, while a wide range of variation is found in P. monogynus, which can have glabrous or sparsely stellate-pubescent to densely stellate-pubescent inflorescences. Physocarpus malvaceus also tends to have petals that are more or less creamy in color and longer stamens, while P. monogynus occasionally has pinkish bracts and petals.
Representative specimens examined. CANADA. British Columbia: 24 mi S of Kootenay Bay along road to Creston, E side of Kootenay Lake, 12 Jun 1953, Calder & Savile 9044 (UC); on a mountain near the lake Edgewood, Lower Arrow Lake, 16 Jun 1960, Ridewood 19 (NY, UC); foothills of the suburbs of Nelson, 8 Sep 1941, Nelson & Nelson 4947 (UC). Alberta: Waterton Lakes National Park, E shore of Waterton Lake, trail to Crypt Lake, 15 Aug 1953, Breitung 17585 (GH).
U.S.A. Idaho: Ada Co., Boise, elev. 2,880 ft., 14 Sep 1911, Clark 326 (GH, UC); Adams Co.,sandy ridge of decomposing granite, 20 mi S of new meadows near Hwy to Weiser, 24 Jun 1946, Hitchcock & Muhlick 13881 (NY); Benewah Co., 5 mi E of Plummer at Heyhorn State Park, elev. 3,700 ft., 12 Jun 1982, Standley 732 (NY); Boise Co., Lowman, on timbered hillsides on edge of meadow, 11 Jun 1947, Christ & Christ 17229 (NY); Bonner Co., forests around Lee Lake, 1 mi E of Coolin along Lee Road, elev. 2,500 ft., 7 Jun 1986, Johnson 1455 (NY); Bonneville Co., Caribou Range, Elk Creek Road, 3.2 km below Jensen Pass, 16 km air distance W-NW of Alpine Junction, elev. 2,165 m, 24 Aug 1997, Holmgren & Holmgren 12963 (NY); Cassia Co., Black Pine Peak areas, Pole Canyon, east-facing slope above Pole spring, 44 airline mi SE of Burley, elev. 8,000 ft., 10 Jun 1969, Holmgren 3512 (NY); Clearwater Co., Clearwater National Forest, Clearwater Mts, between N Fork and Little N Fork Clearwater Rivers on road from Aquarius CG to Smith Ridge, elev. 4,200 ft., 22 Jul 1983, Henderson & Cholewa 6826 (NY); Custer Co., N side of Loon Creek Summit, NW of Estes Mt, NE of Stanley, elev. 8,600 ft., 7 Aug 1986, Atwood 12295 (NY); Elmore Co., above Arrow Rock Lake on road leading to Black Creek, 17 Jun 1947, Hitchcock 15458 (NY, UC); Gem Co., Sage Hen Divide, 9 mi N of Ola, 15 Jun 1941, Christ 12376 (NY); Idaho Co., ca. 1 mi W of Whitebird summit, on edges of Crateagus clumps near creek bank, 20 Jun 1950, Jones 90 (GH, NY); Kootenai Co., Coeur d’Alene, May 1912, Rust 41 (NY); Latah Co., top of Mineral Mt, Skyline drive, elev. 5,020 ft., 25 Aug 1995, Richardson 293 (NY-2 sheets); Lemhi Co., in granite talus, ca. 10 mi S of Gibbonsville, 17 Jun 1944, Hitchcock & Huhlick 9063 (NY); Lewis Co., 4 mi W of Kamiah on Idaho Hwy 64, 2 Jun 1970, Fleak et al. 1485 (NY); Oneida Co., ca. 15 mi ESE of Rockland, elev. 5,000 ft., 5 Jul 1978, Welsh et al. 17293 (NY); Nez Perce Co., Pinnacle Ridge, 14.8 mi S of Grageville, alt, 4,920 ft., 21 Jun 1996, Sondenaa 325 (NY); Shoshone Co., 1 mi below the tower, trail to Little Baldy Lookout, 17 Jul 1946, Witt 1064 (NY, UC); Valley Co., open woods, NE shore of Payette Lake, 6 mi N of McCall, 8 Jul 1953, Baker 10290 (NY); Washtington Co., Schmidt Ranch Road, just S of Adams and Washington county line, elev. 3,600 ft., 13 May 1987, Atwood & Rosentreter 12575 (NY). Montana: Carbon Co., 6 mi SW of Red Lodge, near Hwy to Cooke City, elev. 5,200 ft, 11 Aug 1945, Hitchcock & Muhlick 13476 (GH, NY, UC); Flathead Co., N bank of Flathead River, Belton, 22 Sep 1921, Jack 2444 (A); Gallatin Co., Bridger Canyon, Bozeman, 20 Jul 1898, Blankinship s.n. (UC); Lake Co., SE of McDonald Lake, Mission Range, 12 Jul 1948, Hitchcock 18242 (UC); Lewis and Clark Co., vicinity of Helena, Gates of the Mountains, 8 Aug 1892, Kelsey s.n. (NY); Madison Co., Tobacco Root Mts, 14 Jul 1909, Butler 4266 & 4248 (NY); Missoula Co., foot of Spring Creek Trail, Rattlesnake drainage, 9 Jul 1946, Hitchcock & Muhlick 14592 (NY, UC); Park Co., Livingston, 1 Jul 1901, Scheuber s.n. (NY, UC); Sanders Co., near Graves Creek Falls, N of Thompson Falls, 23 Aug 1976, Kral 58806 (MO); Sweet Grass Co., Wreck Creek, Greycliff, elev. 1,200 m, 28 May – 8 Jun 1912, Eggleston 7891 (A, NY). Nevada: Elko Co., Jarbidge Mts, 3.3 mi N of the town of Jarbidge on the main road to Murphys Hot Spring, elev. 5,800 ft, 28 Jul 1985, Tiehm 10027 (NY). Oregon: Baker Co., Eagle Creek, 5 Jun 1938, Gustafson s.n. (UC); Grant Co., Black Canyon, 12 mi S of Dayville, elev. 3,000 ft., 24 Jul 1953, Cronquist 7607 (NY, UC); Umatilla Co., Gibbon, 4 Jun 1910, Heller 10191 (A); Union Co., on hillsides above the meadow above Morgan Lake, S of La Grande, 16 Jun 1980, Packard & Grimes 80-130 (NY); Wallowa Co., Wallowa Lake campground, 1 mi from Wallowa Lake, 18 Jun 1946, Murphy 42 (NY, UC). Utah: Box Elder Co., Willard Basin Road, 5.2 mi S of Mantua, elev. 2,260 m, 17 Jul 1984, Holmgren & Holmgren 10672 (NY); Cache Co., Cache National Forest, 1 km from junction with Hwy 89, 28 Jul 1994, Walker & Stadler 840 (NY); Carbon Co., Crandall Canyon, above Castlegate, elev. 6,800 ft., 29 Jun 1944, Holmgren & Hansen 3512 (GH, NY, UC); Davis Co., Farmington Canyon, 14 Jul 1902, Pammel & Blackwood 3622 (A-2 sheets, GH); Duchesne Co., Mouth Iron Mine Creek, elev. 9,000 ft., 20 Jun 1939, Hobson s.n. (UC); Garfield Co., 16 mi N of Escalante on Bicknell Road, at Posey Lake, elev. 9,000 ft., 19 Aug 1977, Higgins 11110 (NY); Juab Co., Mt. Nebo, Bear Canyon, elev. 7,200 ft., 4 Jul 1982, Thorne 2077 (NY); Morgan Co., Peterson Creek, Wasatch Mts, 19 Jul 1902, Pammel & Blackwood 3817 (A, GH); Piute Co., Bullion Canyon, Tushar Mts, elev. 7,740 ft., 11 Jul 1984, Taye 2657 (NY); Salt Lake Co., Lambs Canyon, 1 mi S of Parleys Canyon, elev. 6,600 ft., 28 Jun 1982, Neese 11843 (NY); San Juan Co., W slope of LaSal Mts, Upper Brumley Creek, elev. 8,600 ft., 29 Jun 1986, Franklin 3624 (NY); Sanpete Co., near big spring at Fountain Green, 2 Aug 1929, Harris C2955 (UC); Summit Co., Divide between haed of S Fork of Weber River and Hoyt’s Canyon, immediately E of Hoyt’s Peak, elev. 9,500 ft., 11 Jul 1974, Welsh & Murdock 12528 (NY); Tooele Co., S Willow Canyon, near Intake campgound, elev. 1,980 m, 5 Jun 1985, Holmgren & Holmgren 10807 (NY); Utah Co., upper slope of Mt Hollow, Traverse Mts Range, elev. 2,067 m, 1 Jun 1993, Hysell 397 (NY); Wasatch Co., Snake Creek Canyon, ca. 6 mi W of Midway, 12 Jul 1967, Welsh et al. 6320 (NY, UC). Washington: Asotin Co., Fields Spring State Park, 5 mi S of Anatone, elev. 4,000 ft., 27 May 1949 Cronquist 5798 (GH, NY, UC); Ferry Co., hillside along W side of Columbia River 3 mi below mouth of Flat Creek, elev. 1,290 ft., 10 May 1940, Rogers 419 (GH, NY, UC); Okanogan Co., Bonaparte Lake, elev. 3,700 ft., 8 Jul 1921, John et al. 5288 (GH, UC) Spokane Co., hills along roadside to Mt. Spokane, elev. 3,000 ft., 24 Jun 1933, Thompson 9185 (A, GH, NY, UC); Stevens Co., 2.5 mi N of Echo, elev. 3,100 ft., 3 Jun 1992, Laferrière 2351 (A); Walla Walla Co., Waitsburg, 15 May 1897, Horner B172 (GH); Whitman Co., head of moist canyon near junction of US 95 and US 195, 10 mi N of Clarkston, 16 Aug 1941, McVaugh 6397 (A, UC). Wyoming: Lincoln Co., Star Valley Racnch, NE of Thayne plat 2, lot 59, elev. 6,700 ft., 1 Sep 1986, Shaw 4248 (NY); Park Co., Yellowstone National Park, Undine Falls, elev. 6,250 ft., 27 Jun 1979, Lichvar 1874 (NY); Teton Co., Hoback Canyon, Rimrock Ranch Road, 3.5 km air distance SSE of Camp Davis, elev. 2,175 m, 28 Jun 1986, Holmgren & Holmgren 11227 (NY).
5. Physocarpus monogynus (Torr.) J. M. Coult., Contr. U.S. Natl. Herb. 2: 104, 1891 (Fig. 1F); Spiraea monogyna Torr., Ann. Lyceum Nat. Hist. New York 2: 194, 1827; Neillia monogyna (Torr.) Greene, Pittonia 2: 29, 1889; Opulaster monogynus (Torr.) Kuntze, Revis. Gen. Pl. 2: 949, 1891.−TYPE: U.S.A. “Rocky Mts.,” James s.n. (holotype: NY!).
Spiraea opulifolia var. parvifolia A. Gray ex Porter & J. M. Coult., Syn. Fl. Colorado 33, 1874.−TYPE: U.S.A. Colorado: “Rocky Mountain Flora, Lat 39o −41o ,” in 1862, Hall & Harbour 150 (lectotype designated here, NY!; isolectotypes, GH–2 sheets!). Syntype: U.S.A. Colorado: Fremont Co., near Canon city, in 1872, Brandegee s.n. (UC!).
Opulaster glabratus Rydb., Bull. Torrey Bot. Club 31: 559, 1904; Physocarpus glabratus (Rydb.) Rehder, Mitt. Deutsch. Dendrol. Ges. 1915 (24): 221, 1916; Neillia glabrata (Rydb.) Bean, Kew Bull. 1934: 224, 1934.−TYPE: U.S.A. Colorado: West Spanish Peak, alt. 3,000−3,800 m, 9 Jul 1900, Rydberg & Vreeland 6027 (holotype, NY!).
Opulaster hapemanii Rydb., N. Amer. Fl. 22: 244, 1908.−TYPE: U.S.A. Wyoming: Big Horn Mountains, Aug 1892, Hapeman s.n. (holotype, NY!; isotype, UC!).
Shrub to 1.5 m high. Leaves of the flowering branches with blade 1.5−3.8 cm long, 1.7−3.9 cm wide, outline shape ovate, shallowly 3- to deeply 3−5-lobed, the apex obtuse to acute, the base cordate, the margin doubly crenate, the upper surface glabrous, the lower surface glabrous to sparsely pubescent with stellate trichomes; petioles 0.6−1.9 cm long; stipules lanceolate, 4–4.5 mm long, entire, deciduous after anthesis or caducous. Inflorescences more than 10-flowered, glabrous or sparsely pubescent; bracts oblanceolate, 2.3−3 mm long; pedicels 0.8−2 cm long. Hypanthium cupulate to turbinate, 1.8−2.5 mm deep, 3.5−4.2 mm wide at the rim, sparsely to moderately pubescent; sepals ovate, 2.2−3.3 mm long, obtuse to rounded at the apex, sparsely or densely pubescent on both surfaces; petals orbicular, white; stamens 25−30, the longest filaments 2−3.6 mm long; carpels 2, rarely 3, weakly connate to at the base, densely pubescent. Follicles 4−5.5 mm long, not inflated, with spreading beaks; seeds 1−2 per follicle, 1.5−2.5 mm long, yellowish brown to light brown. Chromosome number: n = 9 (Sax, 1931).
Distribution. Western U.S.A., from southern Montana, Wyoming, western South Dakota, central Colorado, New Mexico, western Oklahoma southward northwestern Texas and southeast Arizona (Fig. 5); Douglas fir, white fir, aspen, sprucefir communities; river banks, north-facing slopes, ridges of mountain ranges, often limestone outcrops; 1,980−3,350 m.
This species shows considerable variation in leaf shape, size, and lobing. Normal-sized plants can be confused with P. malvaceus, while smaller plants of P. monogynus are similar to larger plants of P. alternans. Watson (1876) cited Spiraea monogyna and Spiraea opulifolia var. pauciflora as basionyms when he published the name, Neillia torreyi. When Spiraea monogyna was transferred to the genus Neillia the epithet monogyna should have been adopted, therefore the name Neillia torreyi is superfluous and illegitimate (ICN Art. 52.1; McNeill et al., 2012). Syntypes of Spiraea opulifolia var. pauciflora were a mixture of P. malvaceus and P. monogynus (see the nomeclature of P. malvaceus), although the latter species was excluded in the lectotypication (Heller, 1898). Thus, the circumscription of Neillia torreyi by Watson (1876), by citing this taxon as a basionym, included what is now called P. malvaceus and P. monogynus. Because Watson (1876) did not cite any herbarium specimens, it is difficult to identify the original materials. However, there are two Watson specimens collected at Stansbury Island, Utah (collected in 1869) and E Humbolt Mts, Nevada (collected in 1868), both bearing the same collection number (Watson 304). These collection sites match what Watson (1876) described for the distribution of N. torreyi. Interestingly, the former specimen from the Stansbury Island, labeled as Spiraea opulfolia var. pauciflora, is now called P. alternans, while the latter specimen, which does not bear a species name, is very poor material and impossible to identify, but certainly is not P. alternans. Both P. malvaceus and P. monogynus have two (rarely three) carpels, never a single carpel. It is interesting to note that Watson (1876) stated that N. torreyi has one or two carpels, suggesting that his circumscription of N. torreyi included P. alternans in addition to P. malvaceus and P. monogynus.
Representative specimens examined. U.S.A. Arizona: Cochise Co., talus slope of road, right fork of road from Onion Saddle to Rustlers Park, Chiricahua Mts, 9 Jul 1975, Gibson & Horak 3169 (NY); Graham Co., near Taylor Pass, Pinaleno Mts, elev. 7,800 ft, 19 Jun 1917, Shreve 5225 (A). Colorado: Boulder Co., gravelly slope of Spencer Mountain, elev. 9,500 ft., above Eldora, 31 Aug 1962, Jones 34804 (UC); Clear Creek Co., rocky hiilside, Golden-Central city, elev. 1,830 m, 9 Jun 1920, Duthie & Clokey 3792 (A, NY, GH, UC–2 sheets); Custer Co., granitic soil of dry hillside, near San Isabel city, elev. 8,500 ft., 23 Jun 1936, Rollins 1223 (GH, NY); Douglas Co., slopes of Dawson Butte, 22 Jun 1983, Goodman & Lawson 8740 (NY); El Paso Co., Jack Brook, above Minnehaha, lower slopes of Pikes Peak, elev. 8,800 ft., 14 Aug 1924, Bacigalupi 743 (GH, UC); Fremont Co., near Sheep Basin, 8 mi NE of Texas Creek, elev. 9,500 ft., 5 Jul 1954, Harrington 7524 (CS); Gilpin Co., rocky slopes near snow banks above Tolland, elev. 2,800 m, 25 Jun 1926, Palmer 31321 (A); Gunnison Co., Gunnison watershed, Black canyon, elev. 6,800 ft., 8 & 12 Jun 1901, Baker 115 (GH, NY, UC); Huerfano Co., steep bushy hills in ravine, 6 mi E of La Veta Pass on US 160, 8 Jul 1941, McVaugh 5789 (UC); Jefferson Co., 7 mi S of Boulder on Rocky Flats Pediment, 22 Jun 1973, Kunkel & Shultz 225 (CS); Larimer Co., Horsetooth Mountain Park, 1 mi up trail to Horsetooth Rock, elev. 6,000 ft., 17 Sep 1998, Owens & Wen 205 (CS); Las Animas Co., near the top of the north slope of Raton Mesa, elev. 8,000 ft., 11 Jul 1937, Rollins 1856 (GH); Pueblo Co., open ground 10 mi SW of Pueblo, alt 5,300 ft., 28 May 1954, Harrington 7322 (CS); Teller Co., Pikes Peak Forest Reserve, Middle Beaver Creek, elev. 9,800 ft., 24 Jul 1903, Blumer s.n. (GH). Montana: Carbon Co., Beartooth Mts, on the ridge N of the N Fork Grove Creek, ca 6 mi SE of Red Lodge, elev. 7,000 ft, 24 Jun 1986, Lesica 3894 (NY). New Mexico: Bernalillo Co., Sandia Mts, in cool canyon bottom, under white fir and aspen, elev. 9,600 ft, beside La Luz Trail to Sandia Crest from Juan Tabo Canyon, La Cueva Canyon, 18 Jun 1956, Tucker 3091 (UC); Catron Co., Gila National Forest, along Stub Trail Drainage N and NNE of tower on N slope of Bearwallow Mt., elev. 8,120-9,000 ft, Disturbed drainage and conifer forest, locally frequent, 29 Jul 1984, Daniel and Nelson 3592 (NY); Colfax Co., Vicinity of Ute Park, elev. 2,200-2,900 m, 8 Sep 1916, Standley 14539 (A, UC); Grant Co., Black Canyon campground, outer loop road St Hwy 61, 31 mi N of Mimbres, in canyon bottom, Pinyon–Juniper on dry hillsides, on rocky outcrop, elev. 7,100 ft, 18 Jun 1968, Hess 2000 (NY); Lincoln Co., White Mts., Eagle Creek Canyon, alt, ca. 2,500 m, 1 Jul 1984, Worthington 12190 (NY); San Miquel Co., on wooded banks of Pecos River, Pecos, 21 Oct 1939, Brouet and Richards 3321 (A); Santa Fe Co., along Hwy 22 to the Santa Fe Ski Basin, 4 Jul 1973, Higgins 7550 (NY); Sierra Co., 2 mi W Kingston along Percha Creek, elev. 6,200 ft, 30 Jun 1985, Worthington 13334 (NY); Socorro Co., Between Mogollon Ranger Station and Willow Creek (Silver-Willow Creek divide), Gila Forest, elev. 2,100-2,900 m, 29 Jul 1920, Eggleston 16830 (NY); Union Co., Capulin, among rocks at base of Capulin Cone National Monument, 17 Jun 1941, Nelson and Nelson 4691 (UC). South Dakota: Custer Co., Gully below Sylvan Lake, 13 Aug 1950, Brenckle 50163 (NY); Pennington Co., N base of Harney Peak, Black Hills, 7 Aug 1941, Goodman 3320 (NY). Texas: Culberson Co., N Fork of McKittrick Canyon, Guadalupe Mts, 21 Jun 1964, Correll & Hanson 29814 (GH, UC). Wyoming: Albany Co., Canyon of N Fork of Sybille Creek, elev. 6,300 ft., 18 Jun 1957, Porter 7278 (GH, NY, UC); Big Horn Co., Big Horn Range, Shell Creek Canyon, elev. 6,400 ft., 28 Jun 1960, Porter & Poter 8299 (UC); Converse Co., 5 mi NW of Laramie Peak, 4 Jul 1979, Lichvar 1932 (NY); Fremont Co., S flank of the Owl Creek Range, E of Boysen Dam on the Bird’s Eye Pass road, elev. 5,000 ft., 13-15 Jun 1961, Porter & Porter 8592a (GH, UC); Hot Spring Co., Wind River Canyon, 5 mi N of Boysen, elev. 4,400 ft., 16 Jun 1946, Ripley & Barneby 7993 (NY); Johnson Co., Clear Creek 10 mi W of Buffalo, elev. 5,600 ft., 23 Jul 1958, Porter & Porter 7542 (NY, UC); Laramie Co., NW slope of the NE of peak of Twin Mts, 2 mi E of Remount, elev. 7,700 ft., 14 Jul 1975, Nelson & Nelson 1351 (NY); Lincoln Co., Hills E of Afton, elev. 6,500 ft., 24 Jun 1923, Payson & Armstrong 3237 (GH); Park Co., N side of Heart Mt, in ravine between W and E summits, elev. 7,400 ft., 13 Jul 1983, Evert 5424 (NY); Platte Co., Hartville, 20 Jul 1894, Nelson 498 (GH, NY); Sheridan Co., 3 air mi SSW of Dayton, elev. 4,400 ft., 3 Aug 1979, Hartman 10603 (NY).
6. Physocarpus opulifolius (L.) Maxim., Trudy Imp. S.-Peterburgsk. Bot. Sada 6: 220, 1879, nom. cons. (Fig. 1G and H); Spiraea opulifolia L., Sp. Pl. 489, 1753; Neillia opulifolia (L.) W. H. Brewer & S. Watson, Bot. California 1: 171, 1876; Opulaster opulifolia (L.) Kuntze, Revis. Gen. Pl. 2: 949, 1891; Opulaster opulifolius var. typicus C. K. Schneid., Ill. Handb. Laubholzk. 1: 442, 1905.−TYPE: U.S.A. Virginia (holotype, LINN 651.12 (IDC microfiche!)).
Physocarpa opulifolia Raf., New Fl. 3: 73, 1838; Physocarpa glabra Raf., New Fl. 3: 73, 1838.−TYPE: Mts. of East Europe, Sibiria and Oregon (holotype, unknown).
Spiraea opulifolia var. ferruginea Nutt. ex Torr. & A. Gray, Fl. N. Amer. 1: 414, 1840; Neillia opulifolia var. ferruginea (Nutt. ex Torr. & A. Gray) S. Watson, Bibl. Index N. Amer. Bot. 1: 290, 1878; Physocarpus ferrugineus (Nutt. ex Torr. & A. Gray) Daniels, Univ. Missouri Stud. Sci. Ser. 1: 291; 1907.−TYPE: U.S.A. Georgia: Nuttall s.n. (holotype: NY!).
Opulaster intermedius Rydb. in Britton, Man. Fl. N. States 492, 1901; Physocarpus intermedius (Rydb.) C. K. Schneid., Ill. Handb. Laubholzk. 1: 807, 1906; Physocarpus opulifolius (L.) Maxim. var. intermedius (Rydb.) B. L. Rob., Rhodora 10: 32, 1908; Opulaster opulifolius var. intermedius (Rydb.) Farw., Rep. Michigan Acad. Sci. 20: 179, 1918; Neillia intermedia (Rydb.) Bean, Kew Bull. 1934: 224, 1934.−TYPE: U.S.A. South Dakota: Black Hills, on French Creek, 10 miles below Custer, alt. 4,000 ft., 22 Jul 1892, Rydberg 653 (lectotype, NY!, designated by Tiehm, Mem. New York Bot. Gard. 58: 65. 1990; isolectotypes, GH!, UC!).
Opulaster ramaleyi A. Nelson, Bot. Gaz. (Crawforsville) 34: 367, 1902; Physocarpus ramaleyi (A. Nelson) A. Nelson in J. M. Coulter and A. Nelson, Man. Bot. Rocky Mt. 248, 1909.−TYPE: U.S.A. Colorado: near Boulder, 9 Jul 1900, Ramaley 108 (lectotype designated here, RM!). Syntypes: U.S.A. Colorado: near Boulder, 12 Aug 1901, Ramaley 793 (RM!); near Boulder, 20 Aug 1901, Ramaley 874 (RM!). Nelson (1902) cited another specimen (Osterhout 2406) in addition to the three specimens of the Ramaley collection. Because Nelson did not designate the holotype, all of the material would be syntypes. However, he clearly indicated Rameley’s collections were type specimens. Therefore, Osterhout 2406 is a paratype and not a syntype (ICN Art. 9.5).
Physocarpus michiganensis Daniels, Univ. Missouri Stud., Sci. Ser. 1: 291, 1907.−TYPE: U.S.A. Michigan: Kent Co., near McCord’s, Jun 1899, Daniels s.n. (lectotype designated here, UMO!). Syntype: U.S.A. Michigan: Ionia Co., Easton Township, Jun 1896, Daniels s.n. (UMO!).
Physocarpus missouriensis Daniels, Univ. Missouri Stud., Sci. Ser. 1: 291, 1907.−TYPE: U.S.A. Missouri: Boone Co., Columbia, Jun 1904, Daniels s.n. (lectotype designated here, UMO!). Syntypes: U.S.A. Missouri: Boone Co., Columbia, May 1903, Daniels s.n. (UMO!), Columbia, Jul 1904, Aug 1904, Sep 1904, Daniels s.n. (UMO!), Columbia, 20 May 1902, Thom s.n. (UMO!); Jackson Co., 26 May 1891, Bush s.n. (UMO!); St. Louis Co., Jun 1901, Favor s.n. (UMO!). Daniels (1907) included seven syntypes in the original description. One of the original specimens used by Daniels at UMO was labeled as “type material” and two other specimens as “typical material,” while the other four specimens did not bear any such information. These speciemens are considered as syntypes, and the specimen bearing “type material” is here designated as the lectotype.
Opulaster australis Rydb., N. Amer. Fl. 22: 242, 1908; Physocarpus australis (Rydb.) Rehder, J. Arnold Arbor. 1: 256, 1920.−TYPE: U.S.A. North Carolina: Caldwell Co., on the summit of Stone Mountain, 31 Jul 1891, Small & Heller 256 (holotype, NY!; isotypes: US-2 sheets).
Opulaster alabamensis Rydb., N. Amer. Fl. 22: 243, 1908.−TYPE: U.S.A. Alabama: Lee Co., Auburn, 5 Jun 1897, Earle & Baker s.n. (holotype, NY!; isotype, US).
Physocarpus bracteatus (Rydb.) Rehder, Mitt. Deutsch. Dendrol. Ges. 1915 (24): 221, 1916. Opulaster bracteatus Rydb., Bull. Torrey Bot. Club 31: 558, 1904; Neillia bracteata (Rydb.) Bean, Trees Shrubs Brit. Isles. 3: 243, 1933.−TYPE: U.S.A. Colorado: New Windsor, Jul 1898, Osterhout 23 (holotype, NY!).
Physocarpus intermedius f. parvifolius Rehder, J. Arnold Arbor. 2: 128, 1920.−TYPE: U.S.A. Kansas: Shawnee Co., Topeka Capitol Nurseries, Jun 1918, Skinner s.n. (holotype, A!)
Shrub to 3 m high. Leaves of the flowering branches with blade 2.6−7.5 cm long, 2.3−6.6 cm wide, outline shape oval to ovate, shallowly 3-lobed, the apex acute, the base attenuate to rounded, rarely truncate, the margin doubly serrate, the teeth obtuse, the upper and lower surfaces glabrous to densely pubescent with stellate trichomes; petioles 0.6−2.6 cm long; stipules lanceolate, 5 mm long, entire, caducous. Inflorescences more than 10-flowered, glabrous or densely pubescent with stellate trichomes; bracts spatulate, 4−5 mm long; pedicels 1.1−2.7 cm long. Hypanthium cupulate to turbinate, 1.5−2.2 mm deep, 3.5−4.8 mm wide at the rim, densely glabrous or pubescent with stellate trichomes; sepals ovate, 2.2−3 mm long, acute at the apex, glabrous or densely pubescent on both surfaces; petals orbicular, white; stamens 25−30, the longest filaments 4−5 mm long; carpels 3−5, weakly connate to at the base, densely pubescent or glabrous. Follicles 8−12 mm long, highly inflated, with spreading beaks; seeds 2−3 per follicle, 1.5−2 mm long, yellowish brown to light brown. Chromosome number: n = 9 (Sax, 1931); 2n = 18 (Baldwin, 1951).
Distribution. Eastern North America, from Nova Scotia, southward to northern Florida, and westward to South Dakota and central Colorado (Fig. 2); mixed deciduous forests; streams and river banks, lake shores, wooded slopes; 50–2,130 m.
The name P. opulifolius (L.) Maxim. is conserved against P. opulifolius Raf. When Rafinesque (1838) established the genus Physocarpus (originally Physocarpa but now conserved as Physocarpus) he published two new species in the genus. Physocarpus riparius Raf. (no. 668 “P. riparia”) was based on Spiraea opulifolia L. and S. opulifolia var. tomentella DC. The second species, P. opulifolia (L.) Raf. or glabra (no. 669), excluded the type of Spiraea opulifolia L. (Rafinesque, 1838), and should be regarded as a new species in the genus Physocarpus. This publication of his no. 669 makes the epithet opulifolius unavailable in the genus Physocarpus, and could not use the combination “P. opulifolius (L.) Raf.” for his no. 668. Therefore, his no. 668 P. riparius is a legitimate replacement name for S. opulifolia (ICN Art. 11.4). Physocarpus riparius should have been used for S. opulifolia, but has never been used other than in the original publication. Maxim (1879) independently transferred Spiraea opulifolia to the genus Physocarpus, and P. opulifolius (L.) Maxim. has been commonly used (Rehder 1940; Fernald 1970; Gleason and Cronquist, 1991). Proposal to conserve the name P. opulifolius (L.) Maxim. against P. opulifolus Raf. was proposed (Oh, 2004), which was recommended in the General Committee at the 17th International Botanical Congress and adopted in the Vienna Code (Brummitt, 2005) and onwards.
Small (1903) invalidly made a new combination of Opulaster stellatus Rydb. ex Small based on Spiraea opulifolia var. ferruginea. Subsequent combinations, such as Physocarpus stellatus (Rydb. ex Small) Rehder, J. Arnold Arbor. 1: 256, 1920 and Neillia stellata (Rydb. ex Small) Bean, Kew Bull. 1934: 224, 1934 are invalid (ICN Art. 6.10).
Physocarpus opulifolius is fairly common along the river banks and lake shores and distributed widely in eastern North America (Fig. 2). It appears that plants from the southeastern U.S. tend to have stellate-pubescent leaves and inflorescences, which was recognzed as P. intermedius or P. opulifolius var. intermedius. However, the level of pubescence is continuous when more specimens are examined. This treatment does not recognize the pubescent plants as a separate taxon. Nucleotide sequences of the nuclear ITS, the second intron of LEAFY, and chloroplast DNA regions do not show segregation of subdivision in P. opulifolius (Oh and Potter, 2005). Oh and Potter (2005) suggested that P. opulifolius might have been derived by hybridization between P. monogynus and P. capitatus, or their respective ancestors (Fig. 3).
Representative specimens examined. CANADA. Nova Scotia: Lunenberg Co., Lunenberg, 19 Sep 1924, Jack 3515 (A, GH). Ontario: Smooth Rock Falls, 11 Jul 1957, Pease & Wells 38745 (GH); Algoma Dist., Mamainse Pt., 13 Jul 1935, Taylor et al. 2442 (GH); Bruce Co., gravelly shore of Lake Huron, Red Bay, 22 Jun 1933, Breed et al. 148 (GH); Cochrane Dist., Kapuskasing, 20 Jul 1952, Baldwin & Breitung 3323 (GH); Lambton Co., N of Port Franks, 21 Jun 1948, Soper & Dale 3986 (GH); Thunder Bay Dist., Wilson Island, 8 Jul 1937, Hoise et al. 1944 (UC); Welland Co., Point Abino, Lake Erie, 30 Jun 1940, Soper 2034 (GH). Quebec: St. Lawrence River, below Quebec, 9 Jul 1905, Churchill s.n. (GH); Belchasse Co., Beaumont, 14 Jul 1922, Fernald & Pease 25131 (GH).
MEXICO. Coahuila: Cerro El Viejo, Zaragoza, N. L., elev. 2,170 m, 23 Jun 1993, Hinton et al. 22964 (NY).
U.S.A. Alabama: Jefferson Co., along Tarrant Spring Creek, near Killough Springs, 19 Jul 1943, Hughes s.n. (A); Lee Co., Auburn, on dry gravelly bank along streams, 29 Oct 1911, Harbison 736 (A–2 sheets). Arkansas: Benton Co., near Garfield, 20 Oct 1937, Palmer 44454 (NY); Carroll Co., thicket along small rocky stream near Berryville, 7 May 1925, Palmer 27058 (A); Garland Co., 15 mi E of Mount Ida on Hwy 270, 27 Apr 1935, Lodewyks 170 (A); Hot Spring Co., rocky banks of Cove Creek, Magnet cove, 15 Oct 1924, Palmer 26578 (A); Independence Co., 6 mi E of Cushman, 10 Jun 1968, Thomas 8307 (A, NY); Logan Co., Magazine mtn, elev. 2,800 ft, 18 Jun 1938, Demaree 17709 (GH, NY); Marion Co., Cotter, 15 Jun 1914, Palmer 5982 (A, UC); Newton Co, Jasper, 11 Jun 1944, Demaree 25006 (GH); Pope Co., Nogo, 18 May 1933, Merrill 338 (UC); Scott Co., E of Y City on U.S. route 270, 11 May 1991, Wood et al. 4945 (A); Washington Co., Ozark Mts, near West Fork, 3 Jun 1950, Gentry 634 (NY). Colorado: Boulder Co., Mesa trail between Boulder and Eldorado Spring, elev. ca 6,000 ft., outer foothills of Front Range, open Pinus poderosa forest, 28 Jul 1963, Weber 11554 (GH, NY); El Paso Co., neighborhood of Pikes Peak, mouth of Cheyenne Canyon, elev. 6,300 ft., 14 Jul 1896, Bessey s.n. (NY); Larimer Co., Buckhorn Creek, 24 Jun 1897, Osterhout s.n. (GH, NY). Connecticut: Fairfield Co., Norwalk, 20 Jul 1910, Eames 8413 (GH, NEBC); Middlesex Co., Banks of Connecticut River, near East Haddam, 27 Aug 1939, Palmer 44930 (A). Delaware: New Castle Co., bank of Brandywine Creek, Wilmington, without date, Canby s.n. (NY). Florida: Calhoun Co., banks and bottom of small wooded creek, between Ocheesee Pond and Ocheesee Farm, S of Sneads. 9 Oct 1978, Godfrey & Gholson 76691 (GH). Georgia: Bartow Co., on terrace under open tree canopy, along Stamp Creek about 0.25 mi W of junction with McKaskey Creek, elev. 740 ft, 17 Aug 1948, Duncan 8760 (GH); Dade Co., on vertical, SW-facing cliffs in Cloudland Canyon, E of Trenton, elev. 1,300 ft., 8 May 1948, Cronquist 5155 (GH, NY); Early Co., boggy swamp, 2 mi S of Hilton, 26 Mar 1949, Thorne & Muenscher 9245 (GH); Elbert Co., SSW of Hartwell, along Broad River at Georgia route 172, elev. 200 m, 4 May 1982, Spongberg et al. 1731 (A); Floyd Co., N of GA Hwy 172 along Broad River, 3 May 1981, Garland 27 (UC). Illinois: Cook Co., Busse Forest Preserve, 3 mi from Arlington Heights, 13 Jun 1959, Bennett 392 (NY); Hancock Co., Warsaw, 21 May 1963, Chase 17267 (UC); Jo Daviess Co., along the Apple River below dam at Hanover, 26 Sep 1981, Nee 22002 (NY); Knox Co., bank of Spoon River, near Williamsfield, 10 Aug 1949, Chase 10633 (NY); La Salle Co., Starved Rock Park, near Ottawa, 14 JUn 1934, Schumacher s.n. (UC). Indiana: Allen Co., on the bank of Cedar Creek, 31 May 1915, Deam 15830 (A); Delaware Co., near Muncie, 2 Aug 1888, Brady s.n. (GH, UC); Howard Co., along Kokomo Creek, Kokomo, 2 Jul 1941, Ek s.n. (NY); Jasper Co., along Carpenter Creek, 1.3 mi NW of Remington, 27 Jun 1923, Welch 54 (NY); Jennings Co., along Muscatatuck River, 1 mi N of Vernon, 22 May 1942, Friesner 16694 (UC); Knox Co., Wabash River on Government Hill, 8 Jul 1915, Deam 17038 (A); Lagrange Co., wooded, moist bank of Pigeon River, 2 mi NW of Mongo, 31 Aug 1941, Yuncker & Welch 10814 (GH, NY, UC); Monroe Co., Cedar Cliff, 8 May 1887, Blatchley s.n. (NY); Montgomery Co., W bank of Sugar Creek, N of Crawfordsville, 15 May 1930, Bechtel 13138 (NY); Noble Co., near Rome City, 15 Jun 1888, Brady s.n. (UC); Porter Co., Dune Park, 2 Sep 1933, Knowlton s.n. (GH); Pulaski Co., wooded bank of Indian Creek, 4 mi SW of Pulaski, 9 Jun 1923, Deam 38845 (A–2 sheets); Putnam Co., along Walmut Creek, 2 mi NE of Bainbridge, 25 Jun 1911, Grimes 599 (NY–2 sheets); Randolph Co., Cabin Creek raised bog on Rd 1 at Cabin Creek, 6 mi N of Modoc, 9 Jun 1945, Friesner 18920 (GH); Shelby Co., Waldron, May 1934, Rhoades s.n. (A); Starke Co., on the S shore of Bass Lake, 6 Jun 1920, Deam 30901 (NY); Warren Co., moist bank of Pine Creek, 1 mi S of Rainsville, 31 Aug 1916, Deam 21599 (A). Iowa: Allamakee Co., Postville, 8-11 Jul 1904, Pammel et al. s.n. (A); Cedar Co., 4 mi NE of Springdale, 8 Jun 1950, Fay 212 (UC); Clayton Co., along base of wooded bluff S of McGregor, 17 Jun 1923, Shimck s.n. (UC); Dallas Co., along Raccoon River, 18 Aug 1957, Bruggen 3803 (UC); Delaware Co., Backbone State Park, 18 Sep 1923, Shimck s.n. (NY); Dubuque Co., near entrance to White Pine Hollow, 2 mi NW of Luxemburg, 30 Jun 1951, Thorne 10003 (UC); Fayette Co., Fayette, in 1894, Fink s.n. (GH); Hardin Co., Eldora, Aug 1930, Pammel s.n. (A); Muscatine Co., Muscatine, 29 May 1933, Shimck s.n. (NY); Van Buren Co., along river bluffs, near Farmington, 19 May 1929, Palmer 35837 (A); Wapello Co., Ottumwa, 16 Aug 1897, Peters s.n. (GH); Winneshiek Co., in Upper Iowa Valley near Upper Dam, 3 Jul 1933, Tolstead s.n. (UC). Kansas: Cherokee Co., extreme SE corner of the county, 12 May 1940, Horr et al. E300 (GH, UC). Kentucky: Bracken Co., ca. 2 mi N of Brookville on Rt 1159, where it crosses Locust Creek, 21 Jun 1983, Buddell 2206 (NY); Campbell Co., Campbell County Park, ca. 5 mi S of Alexandria on US 27, 19 Sep 1982, Buddeell & Thieret 646 (NY); Fayette Co., Lexington, in 1835, Short s.n. (NY); Pendelton Co., along intermittent stream below dam at Kincaid Lake State Park, ca. 5 mi NE of Falmouth, 14 Jun 1983, Buddell 2141 (NY); Pulaski Co., Cox Bend of S Fork, 24 Apr 1939, Braun 2275 (A); Whitley Co., NE of Jellico, 11 Jul 1937, Smith & Hodgdon 3821 (GH). Maine: Cumberland Co., near Mere Brook, Brunswick, Jul 1914, Furbish s.n. (NEBC); Penobscot Co., near Burr’s Brook, Brewer, 30 Jun 1905, Knight 1860 (GH, NEBC). Maryland: Allegany Co., Cumberland, in 1894, Shriver s.n. (NY); Cecil Co., North East, 20 Jul 1890, Brinton s.n. (NY); Garrett Co., near Crabtree Creek, foot of Backbone mtn, 20 Sep 1980, Hill 9567 (GH, NY); Howard Co., Ellicott City, 3 Aug 1916, Arsène 657 (A); Washington Co., along Bear Branch of Sideling Hill Creek, just N of route 40, 20 Jun 1981, Hill 10354 (NY). Massachusetts: Bristol Co., New Bedford, 27 Jun 1903, Hervey s.n. (NEBC); Essex Co., Georgetown, 14 Jun 1958, Harris 17685b (NEBC); Hampshire Co., Amherst, 12 Jun 1865, Mackie s.n. (NEBC); Middlesex Co., Concord, 23 Sep 1963, Eaton 5632 (NEBC); Norfork Co., Milton, 19 Aug 1990, Churchill s.n. (NEBC). Michigan: Alpena Co., beach of Lake Huron and adjacent beach thickets, Alpena, 2 Aug 1940, Gleason 9905 (NY); Chipperwa Co., Sault Ste. Marie, 22 Jul 1897, Harper & Harper s.n. (A); Emmet Co., Cecil Bay, 11 Jul 1933, Gleason & Gleason 143 (A, GH, NY); Kent Co., near Murray Lake, 27 Aug 1931, Yuncker & Welch 4150 (NY); Keweenaw Co., along shore of Lake Superior, 0.5 mi ESE from Eagle Harbor, 24 Jul 1958, Bennett 382 (NY); Marquette Co., Marquette, 3 Aug 1873, Gillman s.n. (NY); Presque Isle Co., Black Lake, 24 Jul 1927, Gates 14819 (A); Schoolcraft Co., Shore of Lake Michigan, E of Manistique, 17 Sep 1915, Dodge 288 (NY); Washtenaw Co., hills E of Ann Arbor, 5 Oct 1924, Erlanson 833 (NY). Minnesota: Carlton Co., Thompson, 6 Jul 1891, Sandberg 392 (A, NY); Cook Co., S shore of NE promentory of Susie Islands, 13 Aug 1948, Ownbey & Ownbey 1025 (NY, UC); Fillmore Co., Fountain, 13 Jul 1941, Moore 14878 (A, GH); Huston Co., Spring Grove, 3 Jun 1902, Rosendahl 277 (GH); Lake Co., Shore of Lake Superior, Two Harbors, 18 Jul 1940, Gleason 9515 (NY); St. Louis Co., N shore of Lake Superior about 16 mi from Duluth, 18 Jul 1938, Lakela 2564 (A, GH); Wabasha Co., Snake Creek Recreation Area, about 3 mi S of Kellogg, 10 Aug 1986, Smith 12418 (GH); Washington Co., Kinnikinnie Beach, St. Croix River, 2 Oct 1938, Moore & Moore 10614 (NY); Winona Co., summit of bluff, above Lake Winona, 2 Aug 1926, Fassett 3103 (GH, NY). Missouri: Benton Co., near Cole Camp, 10 May 1926, Palmer 30050 (A); Bollinger Co., Patton, 4 May 1932, Kellogg 25865 (A); Carter Co., Current River, Van Buren, 4 Jul 1914, Palmer 6183 (A); Clark Co., Dumas, 7 Jul 1909, Bush 5892 (A, GH, NY); Cooper Co., Moneteau Creek, 8 mi SE of Bunceton, 8 Oct 1938, Steyermark 21775 (UC); Dent Co., Howe’s Mill, 3 Jul 1928, Palmer 34958 (A); Howell Co., Willow Springs, 11 Jul 1941, Palmer 45384 (A, NY); Iron Co., Ironton, 17 May 1923, Palmer 22670 (A); Jasper Co., Jasper, 16 Sep 1908, Palmer 1455 (A–2 sheets); Jefferson Co., SW of Hillsboro, 2.4 km S of Hwy H on Mammouth Road, 21 May 1978, Boufford & Wood 20350 (GH); Jackson Co., bluffs N of Independence, 21 Jun 1895, Mackenzie 613 (NY); Lawrence Co., Red Oak, 1 May 1925, Palmer 26958 (A); Lewis Co., near Monticello, 20 May 1929, Palmer 35906 (A, NY); Lincoln Co., Whiteside, 19 May 1919, Davis s.n. (A); Madison Co., Mine La Motte, 22 May 1926, Palmer 30261 (A, GH); McDonald Co., Noel, 7 Sep 1913, Palmer 4172 (A); Newton Co., Sarcoxie, 14 Jul 1914, Palmer 6266 (A); Ozark Co., near Bakersfield, 7 Oct 1927, Palmer 32840 (A); Pettis Co., Flat Creek, near Sedalia, 9 May 1926, Palmer 30013 (A); Ripley Co., W Fork of Current River, near Ponder, 13 Jul 1933, Palmer 41617 (A, NY); St. Charles Co., 2 mi W of Flint Hill, 31 May 1934, Drouet 1468 (A, GH); St. Francois Co., Bismarck, 25 Jun 1920, Palmer 18047 (A); St. Louis Co., St. Louis, 2 Jun 1875, Eggert s.n. (UC–2 sheets); Shelby Co., near Bethel, 29 Jun 1933, Palmer & Steyermark 40901 (A); Stone Co., James River, Galena, 20 May 1914, Palmer 5678 (A); Taney Co., near Branson, 26 Sep 1923, Palmer 23908 (A). Nebraska: Cherry Co., 26 km E of Valentine, S side of Niobrara River, at beaver pond marshs just E of Cedar Creeks, 7 Jul 1982, Churchill 12172 (NY); Keya Paha Co., 6 mi S of Springview, flood plain of Niobrara River, 15 Jul 1969, Stephens & Brooks 34331 (NY); Rock Co., Niobrara Valley opposite Meadville, 21 Aug 1941, Tolstead s.n. (UC). New Hempshire: Cheshire Co., Walpole, Alogn Great Brook between NH route 12 and River Rd, 25 Jun 1989, Boufford & Wang 25950 (NEBC, NY). New Jersey: Burlington Co., along Crosswicks Creek, Bordentown, 15 Jul 1911, Long 6055 (GH); Camden Co., along Pensauken Creek, Palmyra, 12 May 1915, Long 11916 (GH); Somerset Co., Watchung, 25 Jun 1930, Moldenke 1276 (NY–2 sheets); Sussex Co., bank of Delaware River, below Dingmans Ferry, 20 Jun 1920, Griscom 9597 (GH); Warren Co., Phillipsburg, 15 Aug 1909, MacKenzie 4301 (NY). New York: Albany Co., along water-pipes from Pine Hills, Albany to Rensselaer Lake, 30 Oct 1910, Burnham s.n. (A); Cayuga Co., E shore of Owasco Lake N of Peacock Point, elev. 214 m, 30 Jun 1944, Clausen 6327 (GH, NY); Dutchess Co., Rhinecliff, 23 Jun 1932, House 19255 (GH); Niagara Co., Niagara Falls, 2 Aug 1885, Batchelder 81 (GH); Oneida Co., Utica, 8 Jul 1873, Hunt s.n. (NY); St. Lawrence Co., Canton, 29 Jul 1916, Phelps 1609 (GH); Tompkins Co., Ithaca, 21 Jul 1932, Bean s.n. (GH); Ulster Co., Highland, 11 Jul 1896, Granger s.n. (NY). North Carolina: Ashe Co., near NC 88, 0.6 mi S of Warrensville, 1 Aug 1958, Radford 38535 (UC); Avery Co., Elk River, E of Elk Park, Aug 1953, Barrell s.n. (A–2 sheets); Buncombe Co., Craggy mtn, elev. 5,000 ft., 8 Sep 1897, Biltmore 1282b (A, NY); Clay Co., summit of Stone mtn, 31 Jul 1891, Small & Heller s.n. (UC); Cleveland Co., Nantahala National Forest, 1 km from US 64 on Perry Gap Rd on E side of Buck Creek, elev. 1,090 m, 17 Sep 1982, Spongberg et al. 17223 (A); Davidson Co., 3.5 mi NW of High Rock near NC 8, 16 Jun 1956, Radford 12917 (GH); Guilford Co., bank of small stream about 2 mi SW of Summer, 8 May 1956, Melvin 3325 (NY); Macon Co., 1.5 mi NW of Gneiss, 15 Jun 1946, Radford s.n. (NY); Moore Co., Deep River at Highfalls, 25 Apr 1948, Fox & Godfrey 1367 (GH); Polk Co., road and rich woods along Green River, 13 May 1974, Hardin 13453 (GH); Watauga Co., bank of Watauga River, at Shulls Mill, 8 Sep 1949, Fox & Godfrey 3385 (GH, NY). Ohio: Champaign Co., Dallas Arbor-vitae bog and marl prairie, S of Urbana, 22 Sep 1935, Camp 1003 (NY); Delaware Co., along Olentangy River, 11 Jun 1930, Camp 816 (NY); Franklin Co., Columbus, 1 Oct 1904, Gleason s.n. (NY); Portage Co., Garrettsville, 29 Jul 1905, Webb 782 (GH); Scioto Co., Camp Gordon, Friendship, 31 May 1934, Demaree 10682 (GH, NY, UC–2 sheets). Oklahoma: Delaware Co., wooded calcareous slopes, 3 mi W and 2.5 mi N of state line, W of Siloam Springs, 27 Jul 1949, Waterfall 9198 (GH, NY). Pennsylvania: Allegheny Co., along Pine Creek, 0.5 mi NW of Wildwood, 21 Jun 1939, Henry 734 (UC); Armstrong Co., 2 mi S of Elderton, 5 Jun 1946, Wahl 1810 (GH); Berks Co., ca. 1 mi NE of Inglesmith, 6 Jun 1947, Berkhimer 8566 (UC); Bradford Co., along Susquehanna River, 0.5 mi NW of Wyalusing, 8 Jun 1938, Fosberg 15138 (GH); Bucks Co., Sellersville, in 1883, Fretz s.n. (UC); Butler Co., 1.75 mi S of Ogle, 23 Aug 1938, Fogg 15311 (GH); Centre Co., along Penn Creek, Poe Paddy State Forest Park, 12 Jun 1940, Fogg 16736 (NY); Chester Co., along French Creek, St. Peters, 2 Aug 1924, Pennell 12311 (NY); Columbia Co., along Susquehanna River, Mifflinville, 9 Jun 1938, Fogg 14538 (GH); Cumberland Co., along Conodoguinet Creek, Green Spring, 14 Jun 1940, Adams 5252 (A); Elk Co., wooded alluvial bottom, 1 mi E of Portland Mills, 17 Jun 1940, Wahl 691 (GH); Fayette Co., Big Meadow Run, Farmington, 9 Jul 1931, Core 2931 (NY); Indiana Co., near Home, 14 Jun 1933, Palmer 40336 (A, NY); Jefferson Co., Mill Creek at the Union Township, 18 Jun 1979, Wood & Boufford 4216 (GH, NY); Lancaster Co., near Pleasant Grove, 5 Jun 1901, Heller s.n. (GH); Lehigh Co., Allentown, 11 Jun 1908, Pretz 1242 (NY); Mercer Co., Sharon, 17 May 1886, Spse s.n. (NY); Mifflin Co., along Kishacoquillas Creek, just NW of Yeagertown, 12 Jun 1940, Fender 4341 (UC); Montgomery Co., Unami Creek, 2 mi NE of Sumneytwon, 13 Jul 1935, Ferder 720 (UC); Northampton Co., 2 mi N of Moorestown, 9 Jun 1942, Schaeffer 18375 (GH); Perry Co., roadside, wooded E and W slopes of Trimmers Rock, 1.5 mi E of Newport, 9 Jul 1937, Adams & Adams 3682 (A); Philadelphia Co., Philadelphia, Jun 1949, Alexander s.n. (UC); Pike Co., Millrift, 29-31 May 1916, Wilson s.n. (NY); Snyder Co., Middle Creek, Selingsgrove, 3 Jun 1926, Moldenke 2864 (NY); Washington Co., Washington, Jun 1859, Brewer & Chickering s.n. (NY); Westmoreland Co., along Fourmile Run, ca. 2.5 km SSE of US Hwy 30 on Darlington Rd, 23 Jun 1979, Utech 79-315 (NY). Rhode Island: Rrovidence Co., Providence, 29 Sep 1889, Collins s.n. (NEBC). South Carolina: Abbeville Co., bank along the Savannah River, 0.5 mi S of SC 72 bridge over the Savannah, 6 Sep 1978, Fairey 844 (NY); Anderson Co., bank of Savannah River, just S of mouth of Big Generostee Creek, 20 Sep 1979, Credle 2594 (NY); Oconee Co., Clemson, 20 May 1906, House 2175 (NY). South Dakota: Laurence Co., Rocky ground along Boulder Creek, Boulder Canyon, near Deadwood, 13 Jun 1929, Palmer 37218 (A, NY); Pennington Co., Rock ledge along Box Elder Creek, 12 mi NW of Rapid City, 19 Sep 1969, Stephens 36887 (NY). Tennessee: Claiborne Co., along the Powell River between Brooks Bridge and Burchett Cemetery, 9 May 1985, Wofford 85-15 (GH); Davidson Co., above Cumberland River and railroad, 7 mi WNW of Bordeaux, 16 Apr 1974, Kral 52507 (NY); Hamilton Co., along Tennessee River, near Chattanooga, 20 Apr 1929, Palmer 35443 (A, NY); Knox Co., Tennessee River, Knoxville, Jun 1898, Ruth 270 (NY); Marion Co., SE side of Little Cedar Mtn along Nickajack Reservoir, ca. 2.5 mi NE of Nickajack Dam, 30 May 1985, Webb 5115 (GH); Morgan Co., Clear Fork River, Rugby, 28 May 1934, Underwood 844 (A); Pickett Co., Wolf River just above confluence with Lick Creek, 8 May 1986, Wofford & Ma 86-40 (NY); Roane Co., along Emory River at W edge of Harriman, 2 May 1992, McNeilus 92-189 (GH, NY); Sullivan Co., along Holston River, near brige, 10 mi above Bristol, 19 May 1934, Jennison et al. 3414 (GH); Unicoi Co., along river, Erwin, 18 1934, Underwood & Sharp 3227 (UC). Virginia: Botetourte Co., Flat Top mtn, 25 Jul 1947, Freer 1741 (GH); Charles City Co., James River, SE of Tettington, 19 Sep 1939, Fernald & Long 11342 (GH); Fairfax Co., Homes Run just above Lake Barcroft, 6 Jun 1952, Fosberg 34494 (NY); Fauquier Co., W slope of Bull Run Mts, 1 mi N of Hopewell, 2 Jun 1935, Allard 503 (GH); Frederick Co., Gore 7 Jun 1946, Baldwin 5653 (NY); Montgomery Co., along New River, Berringer mtn, 2 mi E of East Radford, 7 Sep 1935, Adams & Wherry 2268 (A–2 sheets, GH); Rockbridge Co., Balcony Falls, 19 May 1911, Batchelder 2853 (GH); Smyth Co., vicinity of Marion, 6 Jun 1892, Britton et al. s.n. (NY); Warren Co., Front Royal, 19 Apr 1913, Hunnewell 1878 (GH); Wythe Co., Blue Ridge Mts, near Speedwell, 15 Jun 1939, Totten s.n. (NY). Washington D.C.: 10 Jun 1893, Burgess s.n. (NY). West Virginia: Barbour Co., Tygart Valley River Watershed, 9 mi NE of Philippi, 11 Jun 1984, Utech 84-303 (GH); Greenbrier Co., White Sulphur Springs, 4 Sep 1903, MacKenzie 445 (NY); Hampshire Co., Romney, 7 Jun 1946, Baldwin 5654 (NY); Morgan Co., Paw Paw, in 1902, Shear s.n. (UC); Pendleton Co., Snowy mtn, elev. 4,000 – 4,500 ft., 17 Jun 1925, Rydberg 9083 (A, NY, UC); Summers Co., E side of Greenbrier River, 2 mi below Talcott, 7 Sep 1948, Fox 1973 (GH); Tucker Co., bank of the Blackwater River on the Dry Fork, 10 Sep 1904, Moore 2113 (A, GH); Upshur Co., Buckhannon, 10 Jun 1895, Pollock s.n. (A). Wisconsin: Ashland Co., Oak Island, 9 Jul 1938, Fassett et al. 20026 (GH); Brown Co., Cedar swamp, Green Bay, 6 Jul 1878, Schuette s.n. (NY); Buffalo Co., near Fountainn City, 9 Sep 1925, Palmer 28539 (A); Dane Co., Montrose, 17 Jun 1926, Fassett 3158 (GH); Door Co., Garret’s Bay, near Ellison Bay, 15 Sep 1925, Palmer 28786 (A); Marinette Co., between Marinette and Peshtigo, 20 Aug 1933, Grassl 3195 (NY); Racine Co., Racine, 6 Oct 1878, Davis s.n. (GH); Richland Co., 1.5 mi SE of Ithaca, 10 Jun 1912, Lansing 3447 (GH); Sauk Co., Reedsburg, 23 Jun 1932, Haneen 1017 (GH); Waushara Co., Lunch Creek, Wautoma, 7 Jul 1941, Fassett 21345 (A, GH).
Acknowledgements
I am grateful to Daniel Potter, Ellen A. Dean, James A. Doyle, and two anonymous reviewers for invaluable comments and suggestions, to Jean Shepard for managing specimen loans, and to John L. Strother and Kanchi Gandhi for sharing their comments on nomenclature. I also would like to thank the curators and staff of the herbaria listed in Materials and Methods for loans of herbarium specimens and for assistance during visits. This research was supported in part by an NSF Doctoral Dissertation Improvement Grant DEB 0073041 to Daniel Potter and the author, a grant (2015 DNA barcoding for biological resources of Korea) from National Institute of Biological Resources, and the Eco-Innovation Project of the Ministry of Environment, Korea (Grant No. 416-111-005).