한국산 제비꽃속의 화분분류학적 연구
A palynological study of Viola in Korea
Article information

Abstract
한국산 제비꽃속(Viola) 식물 43분류군에 대한 종간 유연관계 분석을 위해 화분학적 연구를 수행하였다. 한국산 제비꽃속의 화분은 모두 단립으로 크기는 소립 또는 중립이었다. 화분의 극축 모양은 각상 또는 원형이었으며 적도면 모양은 장구형, 아장구형, 약장구형으로 구분되었다. 화분의 극성은 모두 등극성이 었으며 발아구는 3–5공구형으로 나타났다. 적도면에서 관찰한 표면무늬는 미세유공-미립상, 미세유공-난선상, 그리고 미세유공-난선-추문-미립상으로 관찰되었다. 이상의 결과에서 한국산 제비꽃속 식물의 화분형태는 매우 유사하여 종간 형질을 기재하는데 어려움이 있었다. 그러나, 야생팬지, 삼색제비꽃, 금강제비꽃, 흰제비꽃, 넓은잎제비꽃, 털낚시제비꽃, 선제비꽃 등 7종류는 발아구 수와 표면무늬, 화분 크기 등에서 형태적으로 유사한 분류군과 뚜렷하게 구분되어 식별형질로 사용이 가능하였다.
Trans Abstract
Palynological studies were conducted to investigate the interspecific relationships among 43 Korean Viola taxa. Pollen of the genus Viola in Korea was monad in terms of the cell unit, and small or medium in size. The shape of the polar view was angular or circular, and the equatorial view was divided into prolate, subprolate, and prolate-spheroidal categories. The pollen polarity was isopolar, and the aperture type was 3–5 colporate. The surface sculptures of the equatorial view were found to be punctate-scabrate, punctate-rugulate, and punctate-rugulate-fossulate-scabrate. Consequently, as the pollen of most Viola taxa have similar morphological characters, it was difficult to construct a palynological description of each species. However, the seven taxa of V. arvensis, V. tricolor, V. diamantiaca, V. patrinii, V. mirabilis, V. grypoceras var. pubescens, and V. raddeana were clearly distinguished with morphologically similar species according to the aperture number, surface patterns and pollen sizes, all of which can be used as useful pollen characters.
서 론
제비꽃과(Violaceae)는 초본 또는 드물게 목본으로 전 세계에 약 22–25속, 800–1,100 여종이 분포한다(Hekking, 1988; Wahlert et al., 2014). 이 중 가장 많은 분류군이 포함된 제비꽃속(Viola)은 대부분 다년생 초본이며 제비꽃과 내에서 가장 진화된 속으로, 580–620여 종이 주로 북반구 온대지역과 열대지역에 분포한다(Clausen, 1964; Ballard et al., 1998; Wahlert et al., 2014). 한국산에 대해서 는 Matsumura (1886)가 졸방제비꽃(V. acuminata) 등 3종을 보고한 것을 시작으로 현재는 약 60종류 이상이 분포하는 것으로 알려져 있다(Korea National Arboretum, 2017).
제비꽃속의 분류체계는 주두의 형태에 따라 5개의 절(section)로 나눈 것이 최초이며(Gingins, 1823), 외부형태학적, 세포학적, 지리적 분포 등 여러 형질을 바탕으로 다양한 분류체계가 제시되었다(Becker, 1925; Clausen, 1927, 1929, 1964; Ballard et al., 1999; Chen, 2006). 한국산에 대해서는 Nakai (1916)가 Gingins (1823)의 분류체계를 따라 3개 절로 정리한 것이 처음이며, 이후 Ishidoya (1929), Kim (1986) 등에 의해 속 내 분류계급이 정리된 바 있다.
제비꽃속의 화분학적 연구는 화분형태, 크기, 발아구(aperture) 수 등의 보고를 바탕으로(Wodehous, 1935; Erdtman, 1952; Gavrilova, 2006), 지역별 제비꽃속의 화분학적 연구(Sun et al., 1994; Shrestha et al., 2005; Mehrvarz et al., 2014), 제비꽃속 화분의 변형능(Mark et al., 2012), 종자형태와의 비교연구(Scoppola and Magrini, 2019), 동일 개체 내 발아구 수가 다른 이형성(heteromorphism) 보고(Dajoz, 1999; Nadot et al., 2000; Lu et al., 2005; Gavrilova, 2006; Furness, 2011; Jang, 2012) 등 다양한 측면에서 연구가 수행되어 왔다.
한국산 제비꽃속 화분 연구 중 Hwang and Kim (1985)은 화분 표면무늬(surface sculpture)를 통해 14분류군을 6개 그룹으로 나누기도 했으며, Whang (2002) 및 Jang et al. (2006)은 종하 분류군을 포함한 근연종을 대상으로 하였으나, 화분 크기, 발아구 수, 표면무늬 등 화분학적 결과만으로는 근연분류군 간 구별이 불가능하다고 보고한 바 있다. 이처럼 한국산 제비꽃속에 대한 화분학적 연구는 대부분 일부 분류군을 대상으로 광학현미경 또는 주사전자현미경 관찰을 통한 형질의 기재만이 보고되어 있어(Lee, 1979; Hwang and Kim, 1985; Chang, 1986; Park et al., 2000, 2001; Whang, 2002; Jang et al., 2006; Jang, 2012; Yoo, 2013), 속내 종간 또는 종하 분류군에 대한 분석은 미흡한 실정이다.
따라서 본 연구에서는 한국산 제비꽃속 전체를 대상으로 화분의 미세구조를 관찰 및 측정하여 분류군 간 유연 관계를 알아보고자 하였다.
재료 및 방법
재료는 국내에 분포하는 60여종(Korea National Arboretum, 2017; National Institute of Biological Resources, 2023) 중 북한에 분포하거나 분류학적 실체가 불분명한 종류를 제외한 36종 5변종 2품종의 총 43분류군을 대상으로 하였다. 시료는 2020년 3월부터 2021년 5월까지 개화기를 중심으로 자생지에서 직접 채집하였고, 일부 개체는 강원대학교 온실에 이식하여 사용하였다. 실험에 사용한 재료의 증거표본은 강원대학교 식물표본관(KWNU)에 보관하였다. 한편 생체 재료 채집이 불가능한 종류는 강원대학교 식물표본관의 석엽표본에서 채취하여 사용하였다(Table 1).

Collection data of Korean Viola used in this study.
채취한 화분은 Erdtman (1952)의 방법을 다소 변형시킨 Livingstone방법(Kim and Lee, 1978)에 따라 초산분해(acetolysis)한 후, Ion Sputter Coater (E-1010, Hitachi Ltd., Tokyo, Japan)를 이용하여 10 nm (15 mA, 20 s) 두께로 Au 코팅하였다. 화분관찰 및 촬영은 시료당 20개씩 선정하여 전계방출주사전자현미경(SUPRA 55VP, Zeiss, Oberkochen, Germany)과 SmartSEM (Zeiss) 소프트웨어를 이용하였다. 촬영 조건은 accelerating voltage 3.0 KV, working distance 2.0–10.3 mm로 하였다. 화분의 입상은 4,000×, 5,000×, 6,000×, 표면무늬는 20,000×로 촬영하였다. 화분형태에 사용한 용어는 Erdtman (1952), Punt et al. (1994), Halbritter et al. (2018)을 기초로 하였으며, 우리말 용어는 Lee (1978)와 Chang (1986)을 따랐다
결 과
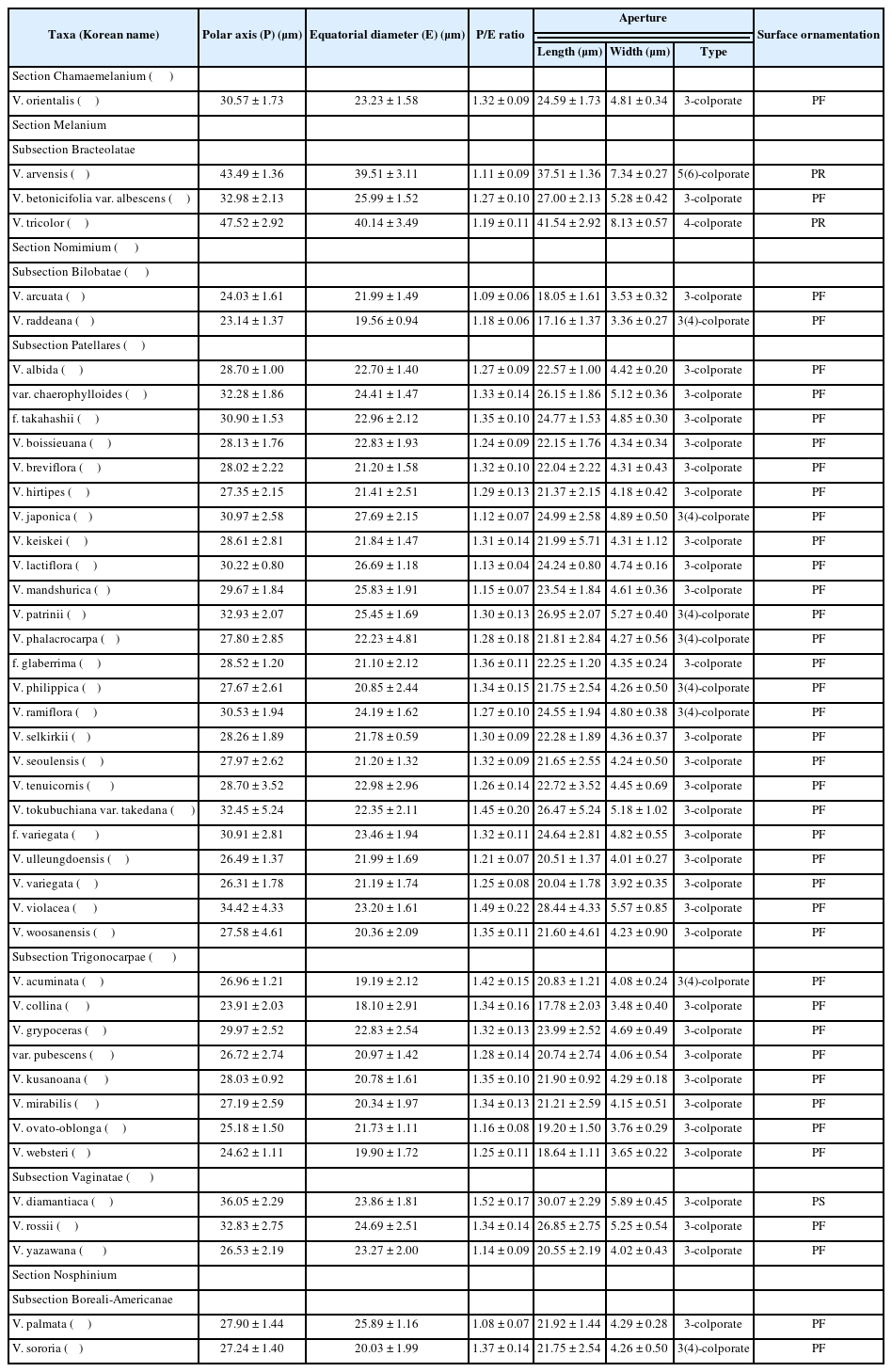
주사전자현미경을 통해 한국산 제비꽃속 43분류군에 대한 화분의 정성적·정량적 형질을 관찰 및 측정한 결과는 Table 2, Figs. 1–8과 같다.
Measurements of pollen grains in Korean Viola.

SEM photographs of pollens in Korean Viola. A. V. orientalis; B. V. arvensis; C. V. betonicifolia var. albescens; D. V. tricolor; E. V. arcuata; F. V. raddeana. 1. equatorial view of pollen; 2. polar view of pollen; 3. sculpturing of pollen (equatorial view). Scale bars: 10 μm.

SEM photographs of pollens in Korean Viola. A. V. albida; B. V. albida var. chaerophylloides; C. V. albida f. takahashii; D. V. boissieuana; E. V. breviflora; F. V. hirtipes. 1. equatorial view of pollen; 2. polar view of pollen; 3. sculpturing of pollen (equatorial view). Scale bars: 10 μm.

SEM photographs of pollens in Korean Viola. A. V. japonica; B. V. keiskei; C. V. lactiflora; D. V. mandshurica; E. V. patrinii; F. V. phalacrocarpa. 1. equatorial view of pollen; 2. polar view of pollen; 3. sculpturing of pollen (equatorial view). Scale bars: 10 μm.

SEM photographs of pollens in Korean Viola. A. V. phalacrocarpa f. glaberrima; B. V. philippica; C. V. ramiflora; D. V. selkirkii; E. V. seoulensis; F. V. tenuicornis. 1. equatorial view of pollen; 2. polar view of pollen; 3. sculpturing of pollen (equatorial view). Scale bars: 10 μm.

SEM photographs of pollens in Korean Viola. A. V. tokubuchiana var. takedana; B. V. tokubuchiana var. takedana f. variegata; C. V. ulleungdoensis; D. V. variegata; E. V. violacea; F. V. woosanensis. 1. equatorial view of pollen; 2. polar view of pollen; 3. sculpturing of pollen (equatorial view). Scale bars: 10 μm.

SEM photographs of pollens in Korean Viola. A. V. acuminata; B. V. collina; C. V. grypoceras; D. V. grypoceras var. pubescens; E. V. kusanoana; F. V. mirabilis. 1. equatorial view of pollen; 2. polar view of pollen; 3. sculpturing of pollen (equatorial view). Scale bars: 10 μm.

SEM photographs of pollens in Korean Viola. A. V. ovato-oblonga; B. V. websteri; C. V. diamantiaca; D. V. rossii; E. V. yazawana; F. V. palmata. 1. equatorial view of pollen; 2. polar view of pollen; 3. sculpturing of pollen (equatorial view). Scale bars: 10 μm.

SEM photographs of pollens in Korean Viola. A. V. sororia. 1. equatorial view of pollen; 2. polar view of pollen; 3. sculpturing of pollen (equatorial view). Scale bars: 10 μm.
화분의크기(pollen size)
한국산 제비꽃속 분류군들의 화분은 단립(monad)으로, 극축의 길이가 10–25 µm인 소립(small)은 콩제비꽃(V. arcuata), 둥근털제비꽃(V. collina), 종지나물(V. sororia), 선제비꽃(V. raddeana), 왕제비꽃(V. websteri)에서 관찰되었으며, 나머지 분류군은 25–50 µm인 중립(medium)이었다. 극축의 길이(polar axis length)는 20.26–52.62 µm 범위였으며, 평균값은 29.55 µm였다. 삼색제비꽃(V. tricolor)이 가장 컸고, 선제비꽃이 가장 작았다. 적도면의 길이(equatorial diameter)는 14.15–46.51 µm로, 평균값은 23.28 µm였으며 삼색제비꽃이 가장 컸고, 둥근털제비꽃이 가장 작았다.
화분입상(pollen shape)
모든 분류군의 화분극성(pollen polarity)은 등극성(isopolar)이였으며, 극면 입상(polar view)은 두 가지 유형으로 야생팬지(V. arvensis)와 삼색제비꽃에서는 각상(angular), 나머지 종류에서는 원형(circular)으로 나타났다. 적도면 입상(equatorial view)은 극축의 길이(P)와 적도면 길이(E)의 비율(P/E)에 따라 3가지로 구분되었다.
장구형(prolate)은 P/E의 값이 1.34–1.99인 형태로, 졸방제비꽃, 단풍제비꽃(V. albida f. takahashii), 둥근털제비꽃, 금강제비꽃(V. diamantiaca), 큰졸방제비꽃(V. kusanoana), 넓은잎제비꽃(V. mirabilis), 종지나물, 민둥제비꽃(V. phalacrocarpa f. glaberrima), 호제비꽃(V. philippica), 고깔제비꽃(V. rossii), 민둥뫼제비꽃(V. tokubuchiana var. takedana), 자주잎제비꽃(V. violacea), 우산제비꽃(V. woosanensis)에서 관찰되었다. P/E의 값이 1.01–1.14인 약장구형(prolate-spheroidal)에는 콩제비꽃, 야생팬지, 왜제비꽃(V. japonica), 흰젖제비꽃(V. lactiflora), 창원제비꽃(V. palmata), 애기금강제비꽃(V. yazawana)이 포함되어 있었다. P/E의 값이 1.15–1.33인 아장구형(subprolate)은 장구형과 약장구형을 제외한 나머지 분류군에서 관찰되었다.
발아구
발아구의 길이(aperture length)는 14.28–46.64 µm 범위였으며, 평균값은 23.52 µm였다. 발아구의 너비(aperture width)는 2.79–9.13 µm로, 평균값은 4.60 µm였다. 발아구의 길이와 너비는 모두 삼색제비꽃이 가장 컸고, 선제비꽃이 가장 작았다. 발아구의 유형(aperture type)은 모두 공구형으로 관찰되었으며, 발아구의 수에 따라 3유형으로 구분되었다. 공구형 발아구가 5개인 5공구형(pentacolporate)은 야생팬지에서 나타났으며, 드물게 6공구형(hexacolporate)도 관찰되었다. 4공구형(tetracolporate)은 삼색제비꽃에서 나타났으며, 3공구형(tricolporate)은 나머지 분류군에서 관찰되었다. 이 중, 졸방제비꽃, 왜제비꽃, 흰제비꽃, 털제비꽃(V. phalacrocarpa), 호제비꽃, 선제비꽃, 가지제비꽃(V. ramiflora)에서는 드물게 4공구형 개체도 나타났다.
표면무늬
모든 분류군의 화분 표면은 공통적으로 작고 둥근 구멍이 있는 미세유공상(punctate)이고, 금강제비꽃을 제외한 나머지 종들은 난선상(rugulate) 내지 추문상(fossulate)의 표면무늬가 발달하였다. 야생팬지와 삼색제비꽃을 제외한 나머지 40분류군에서는 난선돌기 또는 추문돌기 상에 미립돌기(scabrate)가 분포하였다. 이를 바탕으로 화분 표면무늬의 특징은 3유형으로 대별되었다. 미세유공–미립상(punctate–scabrate)은 작고 둥근 구멍이 있고 미립돌기가 발달하는 표면무늬로 금강제비꽃에서 나타났다. 미세유공–난선상(punctate–rugulate)은 작고 둥근 구멍이 있고 난선돌기가 발달하고, 난선돌기 위에는 가로무늬가 있는 특징이 있는데, 야생팬지와 삼색제비꽃에서 관찰되었다. 작고 둥근 구멍이 있고 난선-추문상 또는 추문상 돌기와 더불어 미립돌기가 발달하는 미세유공–난선–추문–미립상(punctate–rugulate–fossulate–scabrate)은 나머지 분류군에서 관찰되었다.
고 찰
한국산 제비꽃속 식물의 화분크기는 소립 또는 중립이며, 3공구형 발아구를 갖는 것으로 보고되어 있다. 한편, 적도면입상은 약단구형 내지 장구형이고 화분 표면무늬는 과립상, 난선상, 망상, 유공상, 추문상, 평활상 등으로 기록되었으나, 동일 분류군에 대한 기재는 연구자별로 차이가있었다(Lee, 1979; Hwang and Kim, 1985; Chang, 1986; Park et al., 2000, 2001; Whang, 2002; Jang et al., 2006; Jang, 2012; Yoo, 2013). 그러나 본 연구에서는 적도면 입상이 약장구형 내지 장구형이고, 표면무늬는 피복층(tectum)에 작은 구멍이 있는 미세유공상이 기본으로 피복층 위에 미립돌기가 있는 미세유공–미립상, 피복층에 난선돌기가 발달한 미세유공–난선상, 그리고 피복층에 난선-추문돌기가 있고 돌기 위에 미립돌기가 발달하는 미세유공–난선–추문–미립상의 3가지 형으로 구분되었다(Figs. 1–8). 선행연구에서 평활상 표면무늬는 광학현미경이나 저배율의 전자현미경에서는 그렇게 판단할 수도 있다고 여겨지나, 미세망상(microreticulate) 또는 망상이라 기재한 것은 명백한 용어 사용의 오류라 판단된다. 더불어 대부분 분류군의 표면무늬는 난선상과 추문상 또는 이 둘이 섞인 난선–추문상으로 두 형을 엄밀히 구분하는 것은 매우 어렵다.
야생팬지와 삼색제비꽃의 발아구는 각각 5공구형과 4공구형을 가지며, 표면무늬는 두 분류군 모두 미세망상이라고 기재된 바 있다(Scoppola and Magrini, 2019). 그러나 본 연구 결과에서는 0.2 µm 정도 두께의 망벽(muri)과, 망강(lumen)의 직경이 0.1 µm 이하로 매우 좁아, 망강 직경이 1 µm 정도일 때 사용하는 미세망상이라는 용어를 적용하는 것에는 동의하지 않고, 미세유공(punctate)이라는 용어를 사용하였다. 더불어 본 연구에서는 20,000×의 고배율하에서 난선돌기에 짧은 횡무늬가 관찰되었다.
털낚시제비꽃(V. grypoceras var. pubescens)의 화분 표면무늬(Fig. 6, D3)는 짧은 난선돌기가 나뭇가지 모양으로 분지되는 독특한 무늬로 나타나, 원변종인 낚시제비꽃(Fig. 6, C3)과 구분되었다. 이는 가는 망상 표면무늬로 보고된(Yoo, 2013) 좀낚시제비꽃(V. grypoceras var. exilis)과 비교해도 뚜렷한 차이가 있었다. 털낚시제비꽃은 외부형태학적으로 거(spur)와 측화피를 제외한 전체에 털이 밀생하는 점으로 미기록 변종으로 보고되었는데(Jang et al., 2009), 본 연구 결과에서도 표면무늬에 큰 차이를 보여 변종 이상의 계급 승격도 가능하다고 판단된다.
한편, 넓은잎제비꽃(Fig. 6, F3)과 흰제비꽃(V. patrinii) (Fig. 3, E3)의 화분표면무늬는 다른 종에서의 난선–추문상 또는 추문상의 돌기보다 넓은 추문돌기를 가져 다른 제비꽃속 화분과 뚜렷한 차이가 있었고, 이는 Halbritter(2016)가 보고한 형태와 일치하였다. 넓은잎제비꽃은 학자에 따라 다른 아절이나 계열로 계급을 조정하거나, 이전하는 등의 견해 차이가 있었는데(Becker, 1925; Clausen, 1927, 1929, 1964; Gershoy, 1934; Kim, 1986), 본 연구 결과는 개화기에는 무경종이나 꽃이 진 후 줄기가 생장하는 형질 등으로 Rostratae아절에서 넓은잎제비꽃만을 별개의 Mirabilis계열로 보는 Chen (2006)의 분류체계를 지지한다.
Vaginatae아절에 포함되는 금강제비꽃, 고깔제비꽃, 애기금강제비꽃 중 금강제비꽃의 화분표면무늬는 미립상으로(Fig. 7, C3), 약한 (난상–)추문상을 보이는 두 종(Figs. 7, D3, E3)과는 차이가 분명하여 다른 형질을 추가 조사하여 종합적으로 판단할 필요가 있다.
민둥뫼제비꽃과 줄민둥뫼제비꽃(V. tokubuchiana var. takedana f. variegata)은 대부분의 형질들이 유사한 형태를 보이고, 잎 앞면의 줄무늬 형질은 고정되지 않은 형질로 두 분류군을 통합해야 한다고 보고되고 있는데(Jung, 2013), 본 화분학적 연구에서도 두 분류군(Fig. 5A, B)의 차이점이 거의 없어 Jung (2013)의 견해를 지지한다.
한편, 뫼제비꽃(V. selkirkii)과 외부형태적으로 유사하지만 부정아가 없고, 개화 후 잎이 커지며 식물체가 크고 저지대에서 자라는 특징으로 신종으로 처리된(Lee et al., 2012) 울릉제비꽃(V. ulleungdoensis)은 엽록체 게놈연구(Go, 2021)에서도 두 분류군이 구분되는 결과를 얻었지만 화분학적 형질에서는 뚜렷한 차이가 없었다(Figs. 4D, 5C).
또한, Nomimum절 Patellares아절에 속하는 알록제비꽃과 자주알록제비꽃(Kim, 1986)은 플라보노이드 성분 분석에서 차이가 없어 동일 분류군으로 처리하거나(Lee and Yook, 1975), 형태적, 미세구조학적, 분자계통학적 특징 모두 매우 유사하고, 잎 앞면의 흰색 줄무늬 유무에 따라서만 구분된다고 보고되고 있다(Jung, 2013). 본 연구에서도 두 분류군(Figs. 4F, 5D)의 화분 특징은 매우 유사하여, 통합 의견을 지지하였다.
이상의 결과를 종합하면, 한국산 제비꽃속 식물의 화분 형질은 많은 분류군이 유사하여 검색표 작성이나 형질 기재에 어려움이 있었다. 그러나 야생팬지, 삼색제비꽃, 금강제비꽃, 흰제비꽃, 넓은잎제비꽃, 털낚시제비꽃, 그리고 선제비꽃 등 7종류는 발아구 수와 표면무늬, 화분 크기 등에서 형태적으로 유사한 분류군과 뚜렷하게 구분되어 식별형질로 사용이 가능하였다.
Acknowledgements
The authors are grateful to anonymous reviewers who provided invaluable comments on the early version of the manuscript. We would like to express our gratitude to Dr. Muyeol Kim, Su-Kil Jang, and Kyeong-Sik Cheon for providing information on the natural habitat and collection specimens.
Notes
CONFLICTS OF INTERST
The authors declare that there are no conflicts of interest.