Carex chromosome numbers from Korean populations (eight taxa)
Article information

Abstract
Carex L. is the most species-rich genus in Korean flora (ca. 180 taxa) as well as worldwide (more than 2,000 species). One of the hypotheses explaining high species diversity in the genus is holocentric chromosomes, which miss constricted centromeres during cell divisions; chromosome fission and fusion events are possible. To provide fundamental biological information, we report the meiotic chromosomes of eight Carex taxa investigated from twelve populations, which include four Ulleung-do Island (oceanic island) populations. The taxa and chromosome numbers observed were C. augustinawiczkii (n = 27II), C. bostrychostigma (n = 22II), C. egena (n = 25II), C. fusanensis (n = 21II), C. japonica (n = 31II, 32II), C. maackii (n = 37II), C. mollicula (n = 32II), and C. pediformis var. pedunculata (n = 36II). Chromosome numbers of C. augustinawiczkii, C. egena, C. fusanensis (a Korean endemic), and C. pediformis var. pedunculata are revealed for the first time. C. japonica from Ulleung-do Island populations exhibits two chromosome numbers, which differ from the chromosome numbers reported from the Korean mainland (peninsula). More chromosome data are required to understand Carex species diversity in Korean flora.
INTRODUCTION
Carex L. is characterized by unisexual flowers on spikes and enclosed bracts of fruits called perigynia and is the most species-rich genus in the temperate zones with more than 2,000 species worldwide, occupying on various environments, sunny, shade, and or damp mountains, riversides, and seashores (Global Carex Group, 2015). In Korean flora, the genus is composed of about 180 species in 42 sections (Park et al., 2016). High species diversity in the genus has been hypothesized by due to holocentric chromosomes.
Holocentric chromosomes miss constricted centromeres during cell divisions, which allow chromosomes to fuse and/ or fission (Hipp et al., 2013). There are some organisms with holocentric chromosomes such as arachnids (spiders and allies), nematode Caenorhabditis elegans, and angiosperms Luzula (Hipp et al., 2013). Among the groups that prossess holocentric chromosome, Carex exhibits high species diversity and the wide ranges of variation in chromosome numbers from n = 6 to n = 66 (Hipp et al., 2009). For that phenomenon, the holocentric chromosome has been hypothesized for being associated with dynamic speciation in Carex (Roalson, 2008; Hipp et al., 2009). Although chromosome numbers are basic and critical information in the genus, 28 out of about 180 Carex taxa in Korea have been investigated (Chung and Chung, 2021; Chung et al., 2023), suggesting further cytological studies are needed.
We here report meiotic chromosome numbers of the following taxa: Carex augustinawiczii Meinsh., C. bostrychostigma Maxim., C. egena H. Lév. & Vaniot, C. fusanensis Ohwi, C. japonica Thunb., C. maackii Maxim., C. mollicula Boott, and C. pediformis var. pedunculata Maxim.
MATERIALS AND METHODS
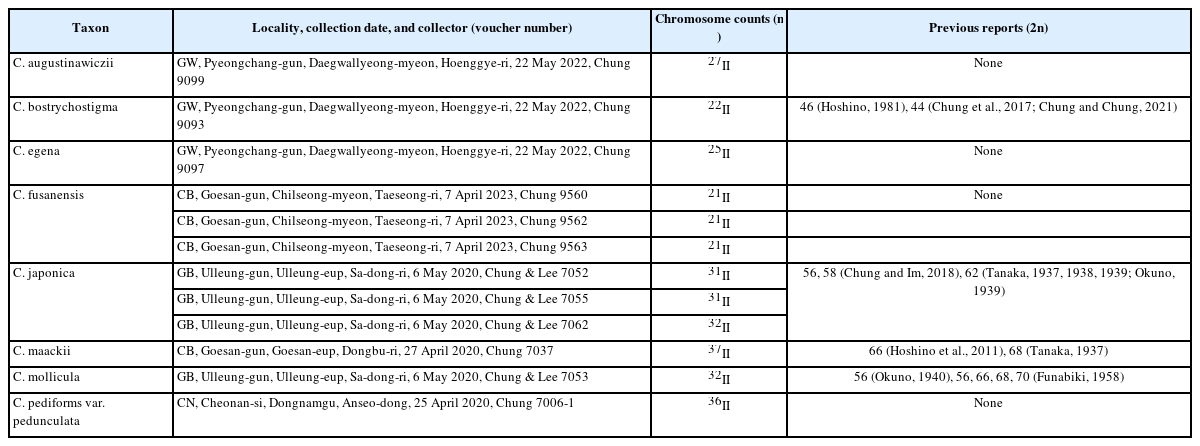
To observe meiotic chromosomes, immature male spikes of Carex were collected and fixed from April 2020 to May 2023. Pollen mother cells of eight Carex taxa from twelve populations were examined. Male spikes were treated in a mixture of methanol, chloroform, and propionic acid at room temperature and then stored in 70% ethanol (Rothrock and Reznicek, 1996). Fixed pollen cells were stained in 1% aceticorcein and squashed. Condensed meiotic chromosomes observed at ×1,000 magnification were drawn, photographed, and analyzed. All counted chromosome numbers were confirmed after at least three meiotic cell observations. A Nikon microscope (Nikon Eclipse 50i, Nikon, Tokyo, Japan) was used for observation, identification followed Hoshino et al. (2011) and Park et al. (2016); and voucher specimens were housed in a herbarium at the Kyungpook National University (KNU).
RESULTS AND DISCUSSION
Chromosomes of eight Carex taxa were confirmed from twelve populations in Korea (Table 1). All chromosomes did not exhibit constricted centromeres during cell divisions and were very small, about 1 μm long (Fig. 1). In the two species examined more than one population, C. fusanensis exhibited a consistent chromosome number whereas C. japonica had two different chromosome numbers.
Chromosome numbers of the Carex taxa investigated in this study and reported in previous studies (CB: Chungbuk, CN: Chungnam, GB: Gyeongbuk, GW: Gangwon).

Photomicrographs of Carex meiotic chromosomes. A. C. augustinawiczkii (n = 27II, Chung 9099). B. C. bostrychostigma (n = 22II, Chung 9093). C. C. egena (n = 25II, Chung 9097). D. C. fusanensis (n = 21II, Chung 9560). E. C. fusanensis (n = 21II, Chung 9562). F. C. fusanensis (n = 21II, Chung 9563). G. C. japonica (n = 31II, Chung & Lee 7052). H. C. japonica (n = 31II, Chung & Lee 7055). I. C. japonica (n = 32II, Chung & Lee 7062). J. C. maackii (n = 37II, Chung 7037). K. C. mollicula (n = 32II, Chung & Lee 7053). L. C. pediformis var. pedunculata (n = 36II, Chung 7006-1). Scale bars = 10 μm.
Carex augustinawiczii Meinsh. 북사초 (n = 27II) (Fig. 1A) – Sect. Racemosae G. Don
Meiotic chromosomes of n = 27II were observed from C. augustinawiczii collected in Pyeongchang-gun, Gangwon-do (Fig. 1A, Table 1). This is the first report of chromosome number for the species. The species habits sunny and wet places such as ravines (glens) in forests, occurring in South Korea, Russia, China, and Japan (Park et al., 2016).
Carex bostrychostigma Maxim. 길뚝사초 (n = 22II) (Fig. 1B) – Sect. Debiles (J. Carey) Ohwi
The meiotic chromosome number for C. bostrychostigma was observed as n = 22II (Table 1 and Fig. 1B). Previous counts from Korean populations are the same (Chung and Im, 2018; Chung and Chung, 2021), but the number from Japan is different (2n = 46) (Hoshino, 1981). The species commonly occurs throughout South Korea and is morphologically and cytologically structured. The species is distributed also in China, Japan, and Russia (Park et al., 2016).
Carex egena H. Lév. & Vaniot 나래사초 (n = 25II) (Fig. 1C) – Sect. Paniceae G. Don
Chromosome number of n = 25II was observed from C. egena collected in Pyeongchang-gun, Gangwon-do (Fig. 1C, Table 1). This is the first chromosome number for the species. The species habits half-sunny and dampish places in forests, occurring in China and Korea (Park et al., 2016; Takahashi et al., 2023). In Korea, the species has been confused with closely related taxa such as C. filipes Franch. & Sav. and/or C. papulosa Boott due to morphological similarity in inflorescences and perigynia. However, C. egena is distinguished from C. filipes by long peduncles of staminate spikes (21–40 mm long) reddish brown staminate scales, and C. papulosa is characterized by green basal sheaths and gradually narrowed long beaks on perigynia (Park et al., 2016; Takahashi et al., 2023). In addition, C. filipes var. oligostachys (Meinsh. ex Maxim.) Kük. is treated as a synonym of C. egena (Takahashi et al., 2023). Somatic chromosome numbers of the C. filipes complex exhibit the wide range of variation from 2n = 40 to 2n = 58, but narrowly distributed species show a consistent chromosome number such as Japanese endemics C. vaniotii H. Lév. (= C. filipes var. vaniotii (H. Lév.) Kük., 2n = 44) and C. rouyana Franch. var. arakiana Ohwi (= C. filipes var. arakiana (Ohwi) Ohwi, 2n = 30) (Hoshino, 1981; Hoshino et al., 2011; Takahashi et al., 2023).
Carex fusanensis Ohwi 부산사초 (n = 21II) (Fig. 1D–F) – Sect. Acrocystis Dumort.
For the first time, a chromosome number for C. fusanensis was revealed as n = 21II (Fig. 1D–F, Table 1). The species has been recognized as C. gifuenesis Franch., an endemic to Japan, but individuals observed in Korea have short ligules (about 1 mm long) and small perigynia (3.0–3.5 mm long), unlike C. gifuenesis which has long ligules (2–3 mm long) and long perigynia (3.5–4.0 mm long) (Hoshino et al., 2011; Park et al., 2016). C. fusanensis is endemic to Korea (Park et al., 2016). C. gifuenensis chromosome number is 2n = 42, the same as the Korean endemic C. fusanensis (Hoshino et al., 2011).
Carex japonica Thunb. 개찌버리사초 (n = 31II, 32II) (Fig. 1G–I) – Sect. Molliculae Ohwi
Three populations of C. japonica in Ulleungdo Island exhibited variation in meiotic chromosome numbers of n = 31II and 32II (Fig. 1G–I, Table 1). The chromosome number of n = 32II is a new number for the species. Previous chromosome reports for the species from Korean populations differ from them and show variation within an individual (n = 28II and 29II) (Chung and Im, 2018). The Ulleungdo Island populations show consistent chromosome numbers within the individuals. The species commonly occurs throughout Korea and is also found in China and Japan (Hoshino et al., 2011; Park et al., 2016). To understand chromosome variation patterns of the species, further investigations covering distribution areas are required.
Carex maackii Maxim. 타래사초 (n = 37II) (Fig. 1J) – Sect. Ovales Kunth
Meiotic chromosome number of n = 37II for C. maackii was counted from a Korean population for the first time (Fig. 1J, Table 1). The chromosome number is a new number for the species. Two different chromosome numbers have been reported (2n = 66 and 68) (Tanaka, 1937; Hoshino et al., 2011). The species is a member of sect. Ovales, which exhibit broad variation in chromosome numbers among and/or within species (Rothrock and Reznicek, 1996; Chung et al., 2011). The species grows on riversides or lakesides and occurs in Korea, China, Japan, and Russia (Park et al., 2016).
Carex mollicula Boott 애기흰사초 (n = 32II) (Fig. 1K) – Sect. Molliculae Ohwi
Meiotic chromosome number of C. mollicula was observed as n = 32II (Fig. 1K, Table 1), which was a new number for the species. Previously, various chromosome numbers for the species (2n = 56, 66, 68, and 70) were reported (Okuno, 1940; Funabiki, 1958). The species is characterized by shorter male (terminal) spikes than female (lateral) spikes and occurs mainly in southern areas of Korea (Park et al., 2016). In addition, the species is also found in China, Japan, and Taiwan (Park et al., 2016).
Carex pediformis var. pedunculata Maxim. 왕그늘사초 (n = 36II) (Fig. 1L) – Sect. Clandestinae G. Don
For the first time, meiotic chromosome number for C. pediformis var. pedunculate was revealed as n = 36II (Fig. 1L, Table 1). This is morphologically similar to C. lanceolata Boott., but long inflorescences, broad leaves, and many lateral spikes (more than 10) characterize C. pediformis var. pedunculata (Park et al., 2016). It is distributed in Korea, China, Mongolia, and Russia (Park et al., 2016).
Chromosome numbers of C. augustinawiczkii, C. egena, C. fusanensis (an endemic species to Korea), and C. pediformis var. pedunculata are revealed for the first time. In addition, C. maackii and C. mollicula from Korean populations are the first reports, that broaden chromosome variation within taxa. Carex japonica from Ulleungdo Island populations exhibits two chromosome numbers, which differ from the chromosome numbers reported from the Korean mainland (peninsula). As in previously reported Carex chromosome numbers, some taxa examined exhibit variation in chromosome numbers. Accumulated Carex chromosome data will contribute to understanding the diversity of the holocentric, species-rich group of plants.
Acknowledgements
We thank Tomomi Masaki (Okayama University of Science) for critical suggestions on chromosome experiments and original data analyses. The present research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2016R1A6A1A05011910) and National Research Foundation (NRF-2021R1I1A3060260).
Notes
CONFLICTS OF INTEREST
The authors declare that there are no conflicts of interest.