Parasenecio pentaphyllaria (Asteraceae: Senecioneae), a new species of Parasenecio from Korea
Article information

Abstract
Parasenecio pentaphylla, a new species of the genus Parasenecio from Mt. Ilwol-san, Ilwol-myeon, Yeongyang-gun, Gyeongsangbuk-do, Korea, is described and illustrated here. In Korea, this species has been recognized as P. adenostyloides, known to be a Japanese endemic species. However, it is clearly distinguished from P. adenostyloides by its involucre shape, the number of phyllaries, the number of florets per capitulum, and the ratio of the tube length to the throat length. A molecular phylogenetic analysis using the Internal Transcribed Spacer region also clearly indicated that P. pentaphyllaria was genetically distinct from P. adenostyloides. Therefore, it is recognized as an independent species, different from P. adenostyloides. The specific epithet “pentaphyllaria” was given based on its characteristic of having five phyllaries.
INTRODUCTION
The genus Parasenecio W. W. Sm. & J. Small (Asteraceae: Senecioneae) comprises approximately 60 species, and it mainly distributed from Northeast Asia to Eastern Europe and the Himalayas (Koyama, 1969; Chen et al., 2011; Kim, 2018). It is characterized by caudate or sagittate anther tail, cylindrical filament collars, and discoid capitula (Smith and Small, 1922; Chen et al., 2011; Xu and Chen, 2016; Chung and Hsu, 2019). In Korea, Parasenecio consists of about 10 taxa (Kim, 2018; Korea National Arboretum, 2023). Korean Parasenecio is characterized by key characteristics such as plant height, number of cauline leaves, shape of petioles, shape of leaves, number of phyllaries, number of florets, shape of florets, and length of achene (Nam, 2004; Chung et al., 2006; Kim, 2018). A taxonomic study was conducted on morphological, cytological, anatomical, and palynological characteristics of seven taxa within Korean Parasenecio (Chung et al., 2006). A molecular phylogenetic study has analyzed the Internal Transcribed Spacer (ITS) region of nrDNA and the trnT-L region of chloroplast DNA for East Asian Parasenecio (Kim, 2001).
Meanwhile, among the taxa of Korean Parasenecio, individuals with angular stem, 3 or 4 cauline leaves, petioles not surrounding the stem, 3-veins or pinnate veins leaves, 3 involucres, and deeply 5-lobed corollas have been identified as P. adenostyloides (Franch. et Sav. ex Maxim.) H. Koyama (Ge-bak-jwi-na-mul, 게박쥐나물). Chung et al. (2006) have reported that the number of phyllaries, a key characteristic, is 5 in all individuals distributed in Korea and 3 in individuals native to Japan. They suggested the need for research to determine whether these differences are due to geographical variations among individuals or represent distinct independent taxa.
In this study, a comparative analysis of morphological and molecular characteristics was conducted on Korean P. adenostyloides and Japanese P. adenostyloides to determine whether they belong to the same taxon.
MATERIALS AND METHODS
Taxon sampling, observation, and measurement
In this study, scientific names and Korean names of Parasenecio based on the recently updated Checklist of Vascular Plants in Korea are used (Korea National Arboretum, 2023). As for taxa with uncertain taxonomic identity or those distributed in North Korea, P. auriculatus (DC.) J. R. Grant var. matsumurana (Nakai) M. Kim (Bak-jwi-na-mul, 박쥐나물), P. hastatus (L.) H. Koyama (Teol-bak-jwi-na-mul, 털박쥐나물), P. hastatus subsp. orientalis (Kitam.) H. Koyama var. ramosus (Maxim.) H. Koyama (Ga-ji-min-bak-jwi, 가지민박쥐), and P. komarovianus (Pojark.) Y. L. Chen (Keunbak- jwi-na-mul, 큰박쥐나물), were excluded; therefore, we collected a total of six taxa distributed in South Korea from August 2021 to September 2022, and these specimens were deposited in ANH herbarium (Table 1). In the Parasenecio genus, plant height, number of cauline leaves, shape of petiole base, leaf shape, number of phyllaries, number of florets, floret shape, style trichome, and synflorescence shape, etc. are recognized as key characteristics (Chung et al., 2006; Chen, 2011; Xu and Chen, 2016; Chung and Hsu, 2019). The herbarium specimens from several herbaria, including JNUB, KB, KH, KIOM, SNUA, and WTFRC, were additionally observed and measured based on these key characteristics. Vegetative characteristics were measured with a Vernier caliper (Digital Caliper, Brütsch/Rüegger Tools Ltd., Urdorf, Switzerland). Reproductive characteristics were observed and measured using a stereo microscope (Olympus SZH10, Tokyo, Japan; IMTcamUSB3.0_ISP6.3, Canada) and an optical microscope (Olympus AX-70, Tokyo, Japan; DP2-BSW, Germany).

Voucher information of the comparative morphological study and molecular phylogenetic analysis, and GenBank accession number in this study.
DNA extraction, PCR amplification, and sequencing
DNA was extracted from about 10–20 mg of dried leaves using DNeasy plant mini-kits (Qiagen Inc., Hilden, Germany). The extracted DNA was quantified using a NanoDrop 2000 (Thermo Fisher Scientific, Wilmington, DE, USA), diluted to a consistent concentration of 15 ng/μL, and stored at −20°C. To amplify the polymerase chain reaction (PCR), we selected the ITS region of nrDNA. The primers used for amplification were ITS1 (5'-TCC GTA GGT GAA CCT GC-3') and ITS4 (5'-TCC TCC GCT TAT TGA TAT GC-3') (White et al., 1990). The total volume of PCR reaction mixture was 20 μL, consisting of 2 μL of genomic DNA, 2 μL of 10 pmole primer pairs, 2 μL of 10× PCR buffer, 4 μL of 5× Q-solution, 0.25 μL of Taq DNA Polymerase (Qiagen, Hilden, Germany), 1.6 μL of 2.5mM dNTP Mixture (Takara, Kusatsu, Japan), and 6.15 μL of deionized water. The PCR cycling conditions were pre-denaturation at 95°C for 2 min, followed by 95°C denaturation for 40 s, annealing at 49–53°C for 40 s and 72°C for 1 min up to 35 cycles, with the final stabilization step carried out at 72°C for 5 min. The amplified PCR products were separated by 1% agarose gel electrophoresis and then purified by commissioning Bioneer (Daejeon, Korea). PCR products were sent to Bioneer for sequencing and sequenced using an ABI 3730XL (Thermo Fisher Scientific, Waltham, MA, USA).
Phylogenetic analysis
The analyzed forward and reverse nucleotide sequences were confirmed for the presence of the target gene using BLAST (Altschul et al., 1997). And assembly was performed using Geneious prime 2022.2.2. (Biomatters Ltd., Auckland, New Zealand). All sequences were aligned using MAFFT (Katoh et al., 2002), and registered in GenBank (Table 1). Syneilesis palmata (Thunb.) Maxim. was selected as an outgroup (Table 1). Maximum parsimony (MP) analysis was conducted using PAUP* 4.0a 169 (Swofford, 2002). All characters were treated as unordered and were weighted equally. Gaps were treated as missing characters. Heuristic searches were replicated 1,000 times additions with treebisection- reconnection (TBR) branch swapping, and accelerated transformation (ACCTRAN). Bootstrap values (Felsenstein, 1985) were calculated from 1,000 replicate analyses by TBR branch swapping to evaluate the support for each clade.
Bayesian inference analysis was performed with MrBayes version 3.2.6 (Huelsenbeck and Ronquist, 2001) plugged in Geneious prime 2022.2.2. (Biomatters Ltd.). The substitution model was selected as GTR+Gamma. Monte Carlo Markov Chain (MCMC) analysis was conducted with 2,000,000 generations, sampling every 500 generations and discarding the 200,000 generations as burn-in. The analyzed phylogenetic tree was constructed using FigTree version 1.4.4. (Rambaut, 2018).
RESULTS AND DISCUSSION
In Korea, Cacalia adenostyloides (Franch. et Sav.) Matsum. or P. adenostyloides (Franch. et Sav. ex Maxim.) H. Koyama are known to be distributed in both Korea and Japan (Lee, 1996a, 1996b; Lee, 2003; Lee, 2007; Kim, 2018; Korea National Arboretum, 2023). In the morphological study, Korean P. adenostyloides has campanulate involucres, usually 5 phyllaries, 5–6 florets in a capitulum, and nearly similar length of corolla tubes and corolla throats (Fig. 1, Table 2). On the other hand, Japanese P. adenostyloides has narrowly cylindric involucres, 3 phyllaries, 3 florets in a capitulum, and corolla tubes that are more than quadruple longer than corolla throats (Fig. 1, Table 2). Results of the molecular phylogenetic analysis, with a high posterior probability (1.00) and bootstrap value (98%), also strongly support that Korean P. adenostyloides is genetically distinct from Japanese P. adenostyloides (Fig. 2). Although it showed no genetic difference in the ITS region compared to P. auriculatus (DC.) J. R. Grant (Gwi-bak-jwi-na-mul, 귀박쥐나물), they were clearly distinguishable based on morphological characteristics (Fig. 2, Table 2). However, it is considered necessary to clarify the genetic relationship with P. auriculatus using different marker regions in the future.
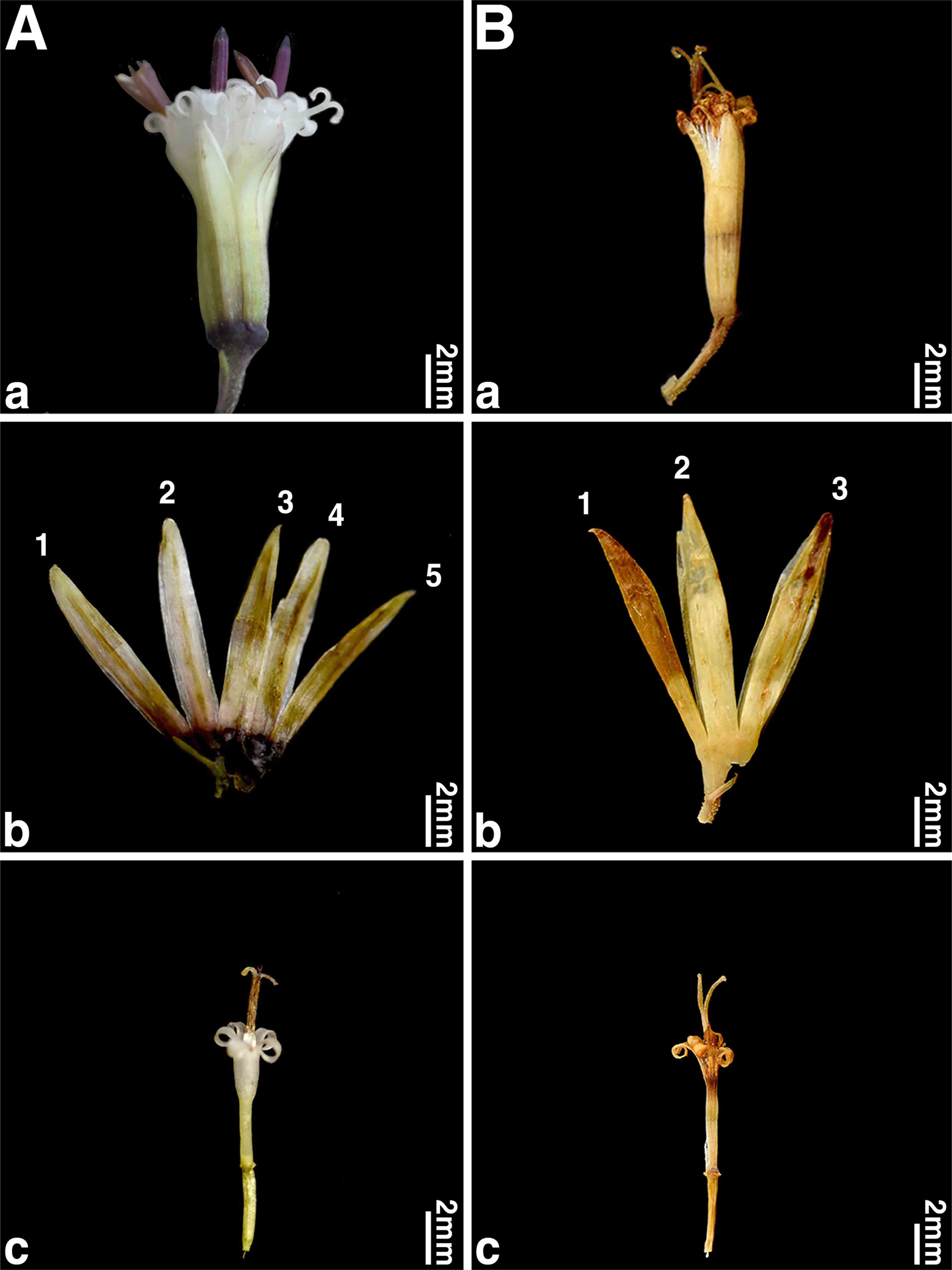
Comparison of morphological characteristics of Korean Parasenecio adenostyloides and Japanese P. adenostyloides. A. Korean P. adenostyloides. B. Japanese P. adenostyloides. a. involucre shape; b. number of phyllaries; c. ratio of corolla tube length to corolla throat length.

Comparison of morphological characteristics of Korean Parasenecio adenostyloides and its related taxa.
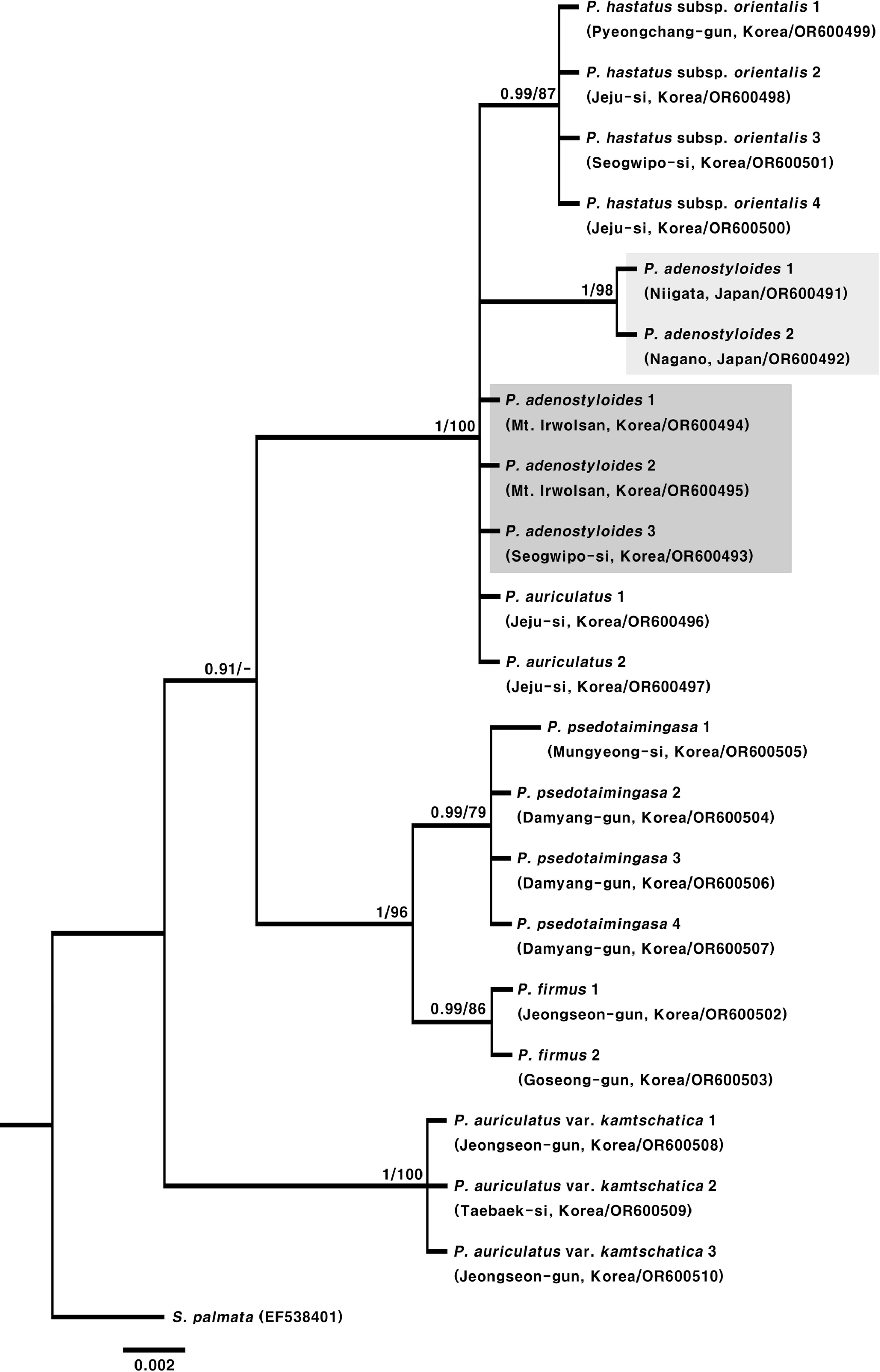
Phylogenetic tree obtained from Bayesian inference analysis based on the Internal Transcribed Spacer (ITS) sequences of Parasenecio. Numbers at nodes show Bayesian posterior probabilities (>0.5)/bootstrap percentages from maximum parsimony analysis, “-” indicates bootstrap value less than 50%.
This species closely resembles P. auriculatus and P. hastatus (L.) H. Koyama subsp. orientalis (Kitam.) H. Koyama (Min-bak-jwi-na-mul, 민박쥐나물) in morphology. Among them, P. auriculatus is characterized by 6–10 cauline leaves and distinct auricle at the base of its leaves. P. hastatus subsp. orientalis is distinguished by 8–24 cauline leaves, winged petioles, triangular-hastate median leaves, 7–8 phyllaries, 11– 13 florets in a capitulum, and corolla throats that are more than twice longer than corolla tubes. These three taxa are clearly distinguished based on these morphological characteristics (Table 2). Other related taxa such as P. auriculatus var. kamtschatica (Kitam.) H. Koyama (Na-rae-bak-jwi-na-mul, 나래박쥐나물), P. firmus (Kom.) Y. L. Chen (Byeong-pung-ssam, 병풍쌈), and P. pseudotaimingasa (Nakai) B. U. Oh (Eo-ri-byeong-pung, 어리병풍) differ in morphological characteristics, including plant height, number of cauline leaves, petioles shape, and median leaves shape (Table 2).
In neighboring countries, taxa similar to Korean P. adenostyloides is P. praetermissus (Pojarkova) Y. L. Chen from China (Chen et al., 2011). Interestingly, P. praetermissus is closely resemble P. auriculatus rather than Korean P. adenostyloides. P. praetermissus is clearly distinguished from Korean P. adenostyloides by its distinctly winged petioles, a petiole base with 1.5–2 cm in width of broad auricle, and a narrowly paniculate synflorescence. In Japan, P. farfarifolia (Sieb. et Zucc.) H. Koyama is morphologically similar to Korean P. adenostyloides. However, this species is characterized by pubescent stems, globose bulbils in axils, deltoid-cordate or pentagonal-cordate cauline leaves, abaxial leaves covered with arachnoid trichomes, and pale-yellow corollas (Koyama, 1995).
As a result of comprehensive research results, the plant that has been identified as P. adenostyloides in Korea is now recognized as a new species.
TAXONOMIC TREATMENT
Parasenecio pentaphyllaria Kwon, Nam & Chung, sp. nov. (Figs. 3, 4).―TYPE: KOREA. Gyeongsangbuk-do, Yeongyang-gun, Mt. Irwolsan, 36°48′21.37″N, 129°05′54.43″E, elev. 1,136m, 12 Aug 2022, M. J. Kwon & G. Y. Chung Ge20220812-001 (holotype: ANH, Fig. 5A; isotype: ANH).

Photographs of Parasenecio pentaphyllaria. A. Habit. B. Rhizome. C. Stem. D. Petiole. E. Median leaf. F. Synflorescence. G. Involucre. H. Capitulum. I. Floret (a. disc flower; b. stamen; c. style). J. Achene.
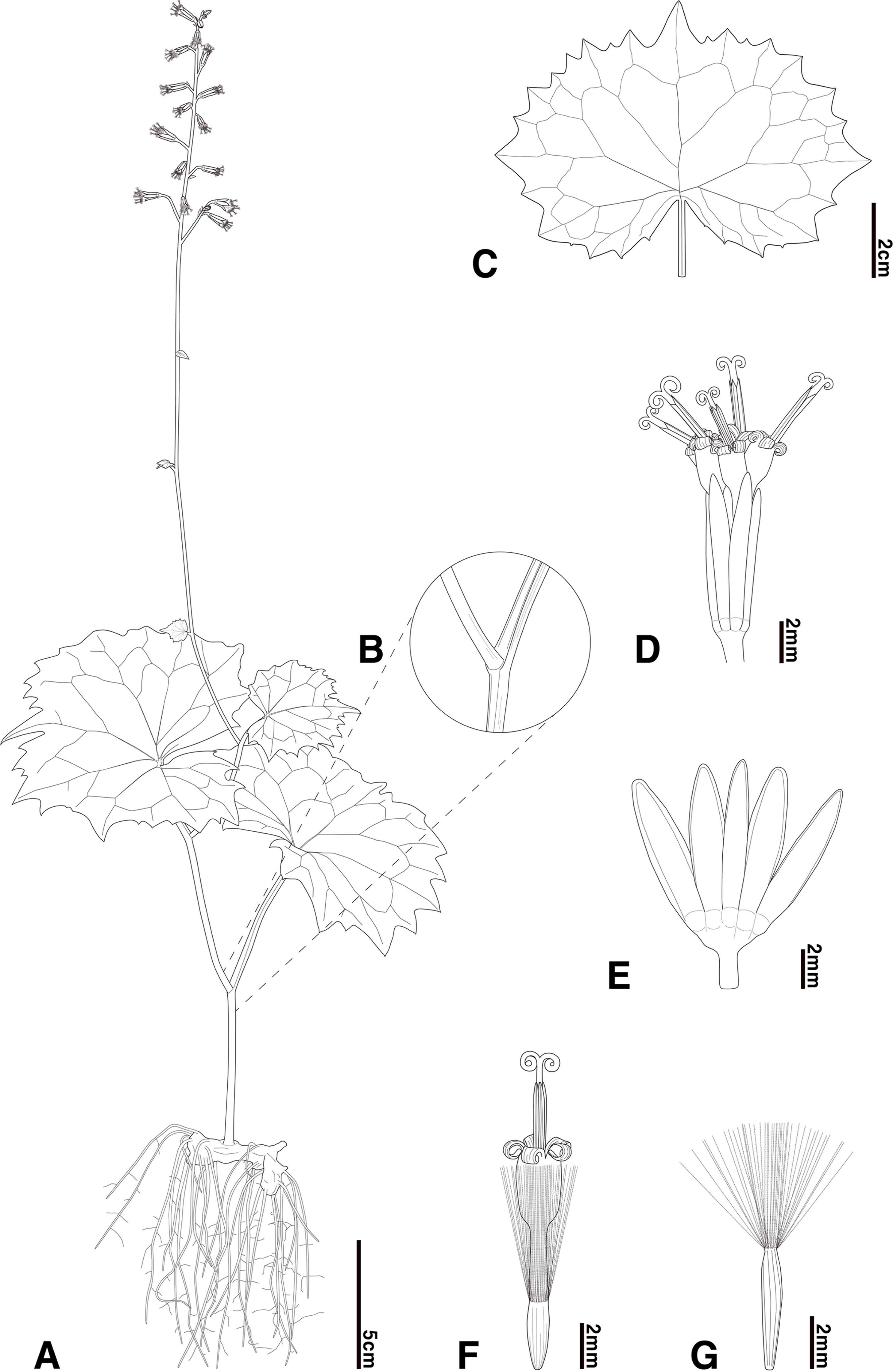
Illustrations of Parasenecio pentaphyllaria. A. Habit. B. Petiole base. C. Median leaf. D. Capitulum. E. Involucre. F. Floret. G. Achene.

Comparison of type specimens in Parasenecio pentaphyllaria and P. adenostyloides. A. Holotype specimen of P. pentaphyllaria [M. J. Kwon & G. Y. Chung Ge20220812-001 (ANH)]. B. Syntype specimen of P. adenostyloides [S. Tschonoski K000814840 (K)].
Korean name: Ir-wol-ge-bak-jwi-na-mul (일월게박쥐나물).
Diagnnosis: Parasenecio pentaphyllaria is morphologically most similar to P. auriculatus and P. adenostyloides among its related taxa. However, this species is clearly distinguished from P. auriculatus by the absence of auricles at the petiole base. Also, this species is distinguished from P. adenostyloides by having campanulate involucres, often 5 phyllaries, 5–6 florets in a capitulum, and equal length of corolla tubes and corolla throats.
Herbs perennial, 23.5–70.1 cm tall. Rhizomes robust, horizontal. Roots numerous fibrous, 2.5–7.2 cm long. Stems erect, zigzaged, shallowly ribbed, 0.8–2.8 mm in diam., light green, green or purple, glabrous. Leaves alternate, simple, 3– 5; petioles 1.5–7 cm × 0.7–1.9 mm, wingless, base not expanded into an auricle, light green or purple, glabrous; lower leaves usually larger than median leaves; median leaves reniform, 5.0–9.9 × 6.5–13.1 cm, base cordate, margin irregular dentate, ciliate, apex mucronulate, abaxial surface green, glabrous, adaxial surface light green, glabrous, veins 5–7 nerved; upper leaves smaller than median leaves. Capitula discoid, 6–54 in an individual, arranged in narrowly raceme or narrowly panicle; peduncles 1.8–8.7 × 0.4–0.6 mm, light green, pubescent; bractioles 1–4, linear to lanceolate, 1.0– 2.9 × 0.3–0.6 mm; involucres campanulate, 7.5–9.0 × 2.3– 3.5 mm; phyllaries 1 series, 5, rarely 4, lanceolate, 6.7–8.3 × 1.2–1.9 mm, light green or purple with white in the upper part, pubescent at apex. Florets bisexual, 5–6; corollas white, glabrous; tubes 2.0–2.9 × 0.5–0.6 mm, throats narrowly cupulate or cupulate, 1.7–2.5 × 1.3–2.2 mm, limbs 4–5-lobed, recurved, 1.5–2.2 mm long; stamen syngenesious, filaments 1.8–3.1 mm long, anthers exserted from corolla, ensiform, 2.3–3.0 mm long, sagittate at base; ovary oblanceolate, 2.5– 3.4 mm long, style 2.7–7.4 mm long, stigma 2-lobed, papillate. Achenes oblanceolate, 3.3–5.9 × 0.7–1.0 mm, brown, glabrous, ribbed. Pappus 3.6–6.6 mm long, white, bristle.
Etymology: The specific epithet “pentaphyllaria” is derived from its characteristic of usually having five phyllaries.
Phenology: Flowering July to August, fruiting September to October.
Distribution and ecology: Korea (Gangwon-do, Gyeongsangbuk-do, and Jeju-do) (Fig. 6). This species inhabits wet and shaded areas at high altitudes.
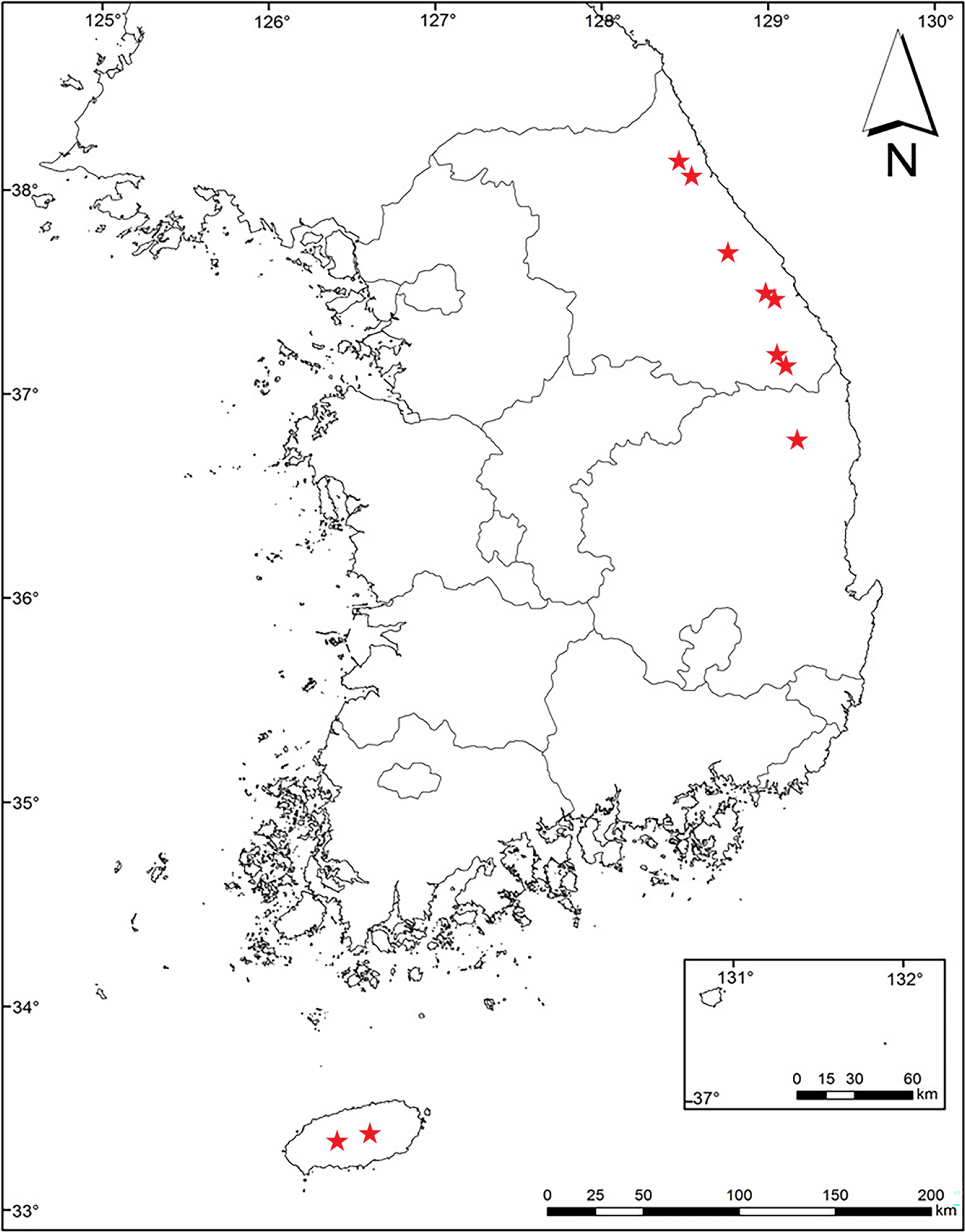
Distribution map of Parasensecio pentaphyllaria in Korea.
Specimens examined: KOREA. Gangwon-do: Inje-gun, Baekdamsa temple, 24 Sep 1966, T.B. Lee & M.Y. Cho 9010 (SNUA); Pyeongchang-gun, Mt. Balwangsan, 16 Aug 2010, K. Heo & D.Y. Jeong 378072-0206 (KB); Samcheok-si, Mt. Dutasan, 14 Aug 2011, B.W. Han NAPI-J20111638 (KB); same locality, 20 Aug 2013, S.C. Kim et al. 20130820001 (KB); Samcheok-si, Mt. Myeonsan, 25 Jul 2012, G.Y. Chung et al. 279133-0090 (KB); Sokcho-si, Mt. Seolaksan, 8 Sep 2006, G.Y. Chung et al. s.n. (ANH0000739); Taebaek-si, Mt. Taebaeksan, 23 Jul 1964, T.B. Lee et al. 3394 (SNUA); Yangyang-gun, Mt. Seolaksan, 25 Jun 1991, J.I. Song s.n. (EWH200508022061, 200508022062); same locality, 13 Sep 2018, J.H. Kim et al. KIMJH18094, 18096 (KB). Gyeongsangbuk-do: Yeongyang-gun, Mt. Irwolsan, 15 Sep 2004, G.Y. Chung et al. s.n. (ANH); same locality, 17 Sep 2004, G.Y. Chung et al. s.n. (ANH040002); same locality, 5 Oct 2004, G.Y. Chung et al. s.n. (ANH); same locality, 2 Sep 2006, M.S. Park s.n. (ANH0000013911, 0000013912); same locality, 20 Sep 2008, G.H. Nam NGH080325 (ANH); same locality, 14 Aug 2015, G.Y. Chung et al. s.n. (KIOM201501014858–201501014860); same locality, 13 Sep 2021, M.J. Kwon & G.Y. Chung Ge210913-1–5 (ANH); same locality, 12 Aug 2022, M.J. Kwon & G.Y. Chung Ge220812-1–14 (ANH); same locality, 29 Sep 2022, M.J. Kwon & G.Y. Chung Ge220929-1–9 (ANH). Jeju-do: Seogwipo-si, Mt. Hallasan, 20 Aug 2009, K.M. Song s.n. (JNUB001042–001045); same locality, 24 Aug 2009, K.M. Song et al. s.n. (JNUB001977); same locality, 19 Aug 2011, G.R. Kim 32302 (WTFRC); Seogwipo-si, Topyeongdong, 6 Sep 2012, M.O. Moon & D.H. Kang 4234 (KB); Seogwipo-si, Hawon-dong, 27 Jul 2022, M.J. Kwon & G.Y. Chung Ge220727-1–4 (ANH); Jeju-si, Eorimok, 7 Jul 1985, D.H. Sin s.n. (JNUB0001454); Jeju-si, Mt. Hallasan, 27 Aug 2005, K.J. Kim et al. 2005-1076 (KB); same locality, 9 Aug 2008, G.Y. Chung et al. s.n. (ANH00005523); Jeju-si, Aewol-eup, 22 Sep 2022, M.J. Kwon & G.Y. Chung Ge220922-1 (ANH).
Key to Parasenecio pentaphyllaria and its related taxa
1. Cauline leaves 1, orbicular-reniform; florets pubescent at the surface.
2. Stems deeply ribbed; cauline leaves 2/3-lobed; involucres cylindric ・・・・・・・・・・・・・・・・・・・・・・・・・・・ P. firmus 병풍쌈
2. Stems shallowly ribbed; cauline leaves 1/2-cleft; involucres narrowly cylindric ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ P. pseudotaimingasa 어리병풍
1. Cauline leaves more than 3, not orbicular-reniform; florets glabrous at the surface.
3. Petioles base auricle.
4. Petioles wingless; cauline leaves reniform; involucres campanulate; corolla throats narrowly cupulate or cupulate, longer than corolla tubes ・・・・・・・・・・・・・・・・・・・・・・・・・・・ ・・・・・・・・・・・・ P. auriculatus var. auriculatus 귀박쥐나물
4. Petioles winged; cauline leaves triangular-hastate; involucres narrowly tubular; corolla throats cylindric, shorter than tubes or same as corolla tubes ・・・・・・・・・・・・ ・・・・・ P. auriculatus var. kamtschatica 나래박쥐나물
3. Petioles base not auricle.
5. Petioles winged; cauline leaves more than 5, triangularhastate ・・・ P. hastatus subsp. orientalis 민박쥐나물
5. Petioles wingless; cauline leaves less than or equal to 5, reniform.
6. Involucres campanulate; phyllaries 5, rarely 4; florets 5–6, corolla tubes same as corolla throats ・ ・・・・・・・・・・・・・・・・・・・・ P. pentaphyllaria 일월게박쥐나물
6. Involucres narrowly cylindric; phyllaries 3; florets 3, corolla tubes more than quadruple longer than corolla throats ・・・・・・・・・・・・・・・・・・・・・・・・ P. adenostyloides
Acknowledgements
This study was carried out with the support of ‘R&D Program for Forest Science Technology (2021400D10-2225DCA02)’ provided by Korea Forest Service (Korea Forestry Promotion Institute).
Notes
CONFLICTS OF INTEREST
The authors declare that there are no conflicts of interest.