/home/virtual/kjpt/journal//../xmls/kjpt-45-2-183.xml
한국산 박주가리아과(광의의 협죽도과)의 화분학적 연구
적 요
한국산 박주가리아과(Asclepiadoideae)에 속하는 4속(Cynanchum, Marsdenia Metaplexis, Tylophora)의 14분류군을 대상으로 해부현미경과 주사전자현미경을 이용하여 화분학적 형질을 관찰 및 측정하였다. 연구 결과, 화분괴(pollinia)의 방향, 세립상의 화분괴 표면무늬는 속을 구분하는 유용한 형질로 나타났다. 한편, 기존에 속 이상의 계급에서 주요 화분학적 형질로 여겨지던 화분괴병(translator-arm)이 붙는 화분괴의 위치는 나도은조롱속(Marsdenia)을 제외하고는 뚜렷이 구분되지 않아 속간 한계를 구분하기에는 어려운 것으로 확인되었다. 또한 왜박주가리속(Tylophora)의 왜박주가리(Tylophora floribunda)에서 유일하게 구상체(corpusculum)의 가장자리가 결각상인 것으로 본 연구에서 처음 관찰되었다. 이외에 화분괴, 화분괴병 및 구상체의 양적 형질에서 크기가 다양하게 나타나 종간 분류에 유용한 것으로 조사되었다.
주요어: 박주가리아과, 화분괴이동체, 화분괴, 화분괴병, 구상체
Abstract
The palynological characters of 14 taxa of 4 genera (Cynanchum, Metaplexis, Tylophora, Marsdenia) in Asclepiadoideae were observed and measured using SM and SEM. The orientation and granulate surface patterns of the pollinia were important characters for delimitating the genera. The position of the attachment of the translator arms to the pollinia, considered to be an important character for recognizing the above genera, was an overlapping character among the genera and can therefore be utilized only as taxonomy at the species level. Also, we found for the first time that only Tylophora floribunda has an incised margin of the corpusculum. The quantitative characters of the pollinia, arms and corpusculum were useful keys for the delimitation of the species.
Keywords: Asclepiadoideae, pollinaria, pollinia, translator-arm, corpusculum
Brown (1810)은 화분 사립체와 화분괴의 유무, 화분괴의 방향과 수 등의 화분학적 형질을 바탕으로 박주가리과(Asclepiadaceae)를 협죽도과(Apocynaceae)로부터 분리하였으나, 최근 다수의 분자생물학적 연구는 두 과가 용담목(Gentianales) 내에서 하나의 단계통군을 형성하는 것을 지지하고 있다( Hallier, 1905; Demeter, 1922; Safwat, 1962; Thorne, 1976, 1992; Struwe et al., 1994; Endress and Bruyns, 2000). 또한 Endress (2004)는 협죽도과 화분을 모식화하여 화분 자루(stalk)와 translator의 발생학적 유연관계를 언급하며 광의의 협죽도과 내 박주가리아과를 인정하기도 했다. 한편, 광의의 협죽도과 내에는 단립의 화분을 갖는 Rauvolfioideae와 Apocynoideae, 사립체의 화분을 갖는 Periplocoideae, 화분괴를 갖는 Secamonoideae 및 박주가리아과의 5개 아과로 구분하는 것이 일반적이다( Endress and Bruyns, 2000; Verhoeven et al., 2003). 그 중 박주가리아과는 172속 약 3,000여 종을 포함하는 가장 큰 아과로서, 온대에서 열대지역까지 널리 분포하고, 화분괴를 가지며 translator가 존재하는 특징으로 다른 아과와 구분된다( Endress and Bruyns, 2000; Endress, 2004).
한국산 광의의 협죽도과 내에는 단립의 화분을 갖는 Rauvolfioideae ( Amsonia)와 Apocynoideae ( Apocynum, Nerium, Trachelospermum, Trachomitum)의 2개 아과 내에 5개 속이 포함되며, 화분괴를 갖는 박주가리아과에는 5속( Cynanchum, Hoya, Marsdenia, Metaplexis 및 Tylophora)의 20분류군이 전국적으로 분포한다고 알려져 있다( Lee, 1996; Lee, 2003, Lee, 2006; Lee, 2007). 박주가리아과는 나도은조롱( M. tomentosa Morren & Decne.)과 호야( Hoya carnosa (L.f.) R.Br.)가 속하는 나도은조롱족(Marsdenieae)과 나머지 분류군을 포함하는 박주가리족(Asclepiadeae)의 2족으로 구성되며, 이 중에는 재배 및 관상용 식물인 넓은잎큰조롱( C. auriculatum Royle ex Wight)과 호야를 비롯하여, 남한에는 분포하지 않는 것으로 알려진 좀박주가리( C. chinense R.Brown), 양반풀( C. thesioides (Freyn) K.Schum.), 자주박주가리( C. purpureum (Pall.) K.Schum.), 세포큰조롱( C. volubile (Maxim.) Hemsl.) 등이 포함되어 있다( Lee, 1996; Lee, 2003, Lee, 2006; Lee, 2007). 이들 분류군 중 최근에 중국 동북부 지역과 북한의 서쪽 해안가에 주로 분포한다고 알려진 좀박주가리와 북한의 강원도 세포지역에서 처음 발견되어 국명이 붙여진 세포큰조롱은 각각 인천과 경북, 제주 지역에서 분포가 확인되고 있다. 따라서 이와 관련된 분포학적 재고 뿐 만 아니라 현재까지 국내에서 이루어진 바 없는 박주가리아과의 형태학적 및 화분학적 유연 관계 등의 종합적인 분류학적 연구가 필요한 것으로 판단된다. 앞서 언급한 바와 같이 화분학적 형질은 협죽도과의 분류학적 한계설정에 가장 중요한 형질로 여겨지고 있으나, 한국산 식물을 대상으로 한 구체적이고 체계적인 연구는 미흡한 실정으로 정확한 화분 형태를 통한 분류군간 유연 관계를 파악하는 것이 요구된다. 따라서 본 연구는 해부현미경과 주사전자현미경을 통하여 화분학적 구조와 형태를 관찰하고 기재하여 본 아과의 화분학적 형질의 중요성과 그들의 분류학적 유연관계를 밝히는 데 목적이 있다.
재료 및 방법
한국산 박주가리아과에 포함되는 5속 20분류군 중 채집 가능한 4속 14분류군의 화분괴이동체는 2011년 5월부터 2014년 8월까지 야외에서 채집된 액침표본(FAA 또는 70% 에탄올에 고정)으로부터 채취하였다( Appendix). 시료는 분류군당 20개씩 측정하였으며, 슬라이드 글라스 위에 올려 해부현미경(SM; Olympus SZH-10, Japan)으로 관찰, 측정 및 촬영(Olympus DP2-BSW, Germany) 하였다. 주사전자현미경 관찰을 위해서는 시료를 stub위에 올린 뒤 실온에서 완전히 건조시키고, ion sputter (Cressington sputter coater 108, USA)를 이용하여 30 mA에서 1분간 gold coating 후, 주사전자현미경(SEM; Jeol JSM-6300, Japan)을 이용하여 15 kV에서 관찰 및 촬영하였다. 화분 형태의 기재에 사용된 용어는 Bookman (1981)과 Singer et al. (2008)를 따랐으며( Table 1), 우리말 용어는 Chang (1986) 등을 따랐다.
결 과
한국산 백미꽃속속에 속하는 11분류군의 화분괴이동체 형태를 관찰한 결과, 화분괴의 평균 길이는 147−420 μm, 평균 너비는 67−276 μm로( Table 1), 크기가 가장 큰 것은 민백미꽃( C. acuminatifolium (Decne.) Hemsl.)( Fig. 2A), 가장 작은 것은 선백미꽃( C. inamoenum (Maxim.) Loes.)( Fig. 2E)으로 나타났다. 화분괴의 형태는 좁은 타원체형(narrowly ellipsoid), 타원체형(ellipsoid), 넓은 타원체형(broadly ellipsoid)으로 관찰되었으며, 화분괴는 모든 분류군에서 직립하였고, 화분괴의 표면은 평활상(psilate), 아망상(sub-reticulate) 또는 망상(reticulate)의 표면무늬를 갖는 것으로 확인되었다. 화분괴병의 평균 길이는 66−187 μm로 가장 짧은 것은 세포큰조롱( C. volubile (Maxim.) Hemsl., Fig. 2J), 가장 긴 것은 민백미꽃이었다( Table 1, Fig. 2A). 화분괴병은 덩굴박주가리( C. nipponicum Matsum.)와 흑박주가리( C. nipponicum var. glabrum (Nakai) H. Hara)의 2분류군만 화분괴의 중부(middel)에 붙었으며( Fig. 2G- H), 나머지 분류군은 모두 화분괴의 상부(apical) 또는 아상부(sub-apical)에 붙는 것으로 나타났다. 구상체의 평균 길이는 188−407 μm, 평균 너비는 73−260 μm로 가장 큰 것은 민백미꽃( Fig. 2A), 가장 작은 것은 선백미꽃이었다( Table 1, Fig. 2E). 구상체의 형태는 좁은 타원체형, 타원체형, 난형(ovoid), 넓은 난형(broadly ovoid), 도란형(obovate)의 형태가 관찰되었으며, 가장자리는 모두 매끈했다( Fig. 2A- K).
나도은조롱속(Marsdenia)의 화분괴이동체 형태(Fig. 2L, Fig. 3L)
본 속의 국내 분포하는 유일한 종인 나도은조롱( M. tomentosa Morren & Decne.) 화분괴의 길이는 500−633 μm, 너비는 200−233 μm인 타원형이며( Table 1), 직립하고, 화분괴의 표면은 아망상이다. 화분괴병의 길이는 133−217 μm이며( Table 1), 화분괴병은 화분괴의 기부(basal)에 붙는 것으로 나타났다. 구상체는 길이가 300−367 μm, 너비가 167−200 μm인 도란형으로 가장자리는 매끈했다( Table 1).
박주가리속(Metaplexis)의 화분괴이동체 형태(Fig. 2M, Fig. 3M)
본 속 가운데 국내에 분포하는 유일한 종인 박주가리( M. japonica (Thunb.) Makino)의 화분괴의 길이는 467−567 μm, 너비는 233−300 μm인 타원체형으로 직립하고( Table 1), 화분괴의 표면은 평활상이었다. 화분괴병의 길이는 133−183 μm( Table 1), 화분괴에 붙는 화분괴병의 위치는 화분괴의 아상부(sub-apical)로 확인되었다. 구상체는 길이가 400−467 μm, 너비가 133−200 μm인 좁은 타원체형으로 ( Table 1), 가장자리는 매끈했다.
왜박주가리속(Tylophora)의 화분괴이동체 형태(Fig. 2N, Fig. 3N)
본 속의 국내 분포하는 유일한 종인 왜박주가리( T. floribunda Miq.)의 화분괴는 길이가 63−125 μm, 너비가 125−150 μm인 타원형으로 한국산 박주가리아과 중 화분괴의 크기가 가장 작았다( Table 1). 화분괴는 가로(transverse) 방향으로 달리며, 화분괴병에 매달린 형태(pendulous)이다. 화분괴의 표면은 세립상(granulate)으로 나타났다. 화분괴병의 길이는 13−38 μm로 본 연구에서 취급된 분류군 중 가장 짧은 것으로 확인되었다( Table 1). 구상체는 길이가 38−75 μm, 너비가 38−75 μm인 도란형이며( Table 1), 가장자리는 결각상(incised)으로 나타났다.
고 찰
광의의 협죽도과(Apocynaceae s.l.) 내에서는 크게 단립의 화분, translator (구상체와 화분괴병을 통칭)에 싸여있는 많은 사립체를 갖는 화분 및 4개 또는 2개의 화분괴를 갖는 화분괴이동체의 특징으로 아과를 구분할 수 있으며( Endress and Bruyns, 2000; Verhoeven et al., 2003), 광의의 협죽도과내 화분학적 진화경향성은 단립에서 화분괴의 형태로 진화했다고 보고된 바 있다( Endress, 2004; Harder and Johnson, 2008). 그 중 박주가리아과(Asclepiadoideae) 내에서 화분괴의 방향은 족내의 분류계급을 결정하는 중요한 형질 중 하나로 여겨져 왔다( Brown, 1810; Swarupanandan et al., 1996; Civeyrel et al., 1998; Shah and Ahmad, 2014). 박주가리아과 중 국내에는 분포하는 족은 2개이며, 이들은 나도은조롱( Marsdenia tomentosa Morren & Decne.)을 포함하는 나도은조롱족(Marsdenieae)과 나머지 분류군이 속하는 박주가리족(Asclepiadeae)의 2개 족으로 구분된다. 한국산을 대상으로 화분괴의 방향을 관찰한 결과( Table 1, Fig. 2), 나도은조롱족은 직립하는 화분괴를 가지며( Fig. 2L), 박주가리족은 직립하는 화분괴와 가로 방향의 화분괴를 둘 다 갖는 것으로 나타나 화분괴의 방향으로 두 족을 명확히 구분하는 것은 어려운 것으로 나타났다( Fig. 2A- N). Shah and Ahmad (2014)은 나도은조롱족 식물 중 가로 방향의 화분괴를 갖는 호야( Hoya carnosa (L.f.) R.Br.)의 경우와 같이 화분괴의 방향으로 족을 구분할 때 예외의 경우가 있다고 언급한 바 있다. 국내에는 백미꽃속( Cynanchum)을 제외하고 3속에 각 1종씩 분포하고 있어 족 단위에서 유연관계를 설명하기에는 어려우나, 왜박주가리속( Tylophora)은 화분괴가 가로 방향으로 달리는 점에서 다른 속과 뚜렷이 구분되었다. 따라서 한국산의 경우 화분괴의 방향은 속을 구분하기에는 유용한 형질로 판단된다. 화분괴 표면무늬는 왜박주가리속에만 세립상의 형태로 나타나 다른 속들과 뚜렷이 구분되었으며( Fig. 3N), 나머지 속에서는 모두 평활상, 망상 또는 아망상의 형태로 나타났다( Fig. 3A- M). 하지만 평활상, 망상 및 아망상의 형태 사이에는 다소 연속적인 변이가 존재하여, 종간 구분에는 어려움이 있는 것으로 판단된다. Meve et al. (2002)은 필리핀산 Tylophora parviflora Wight의 화분괴 표면무늬를 우상돌기(verrucose surface)라고 보고했으며, Tseng et al. (2011) 역시 Tylophora sui Y.H. Tseng & C.T. Chao의 화분괴 표면무늬를 SEM으로 촬영한 결과, 세립상의 표면무늬를 갖는 것으로 보고하고 있다. 따라서 기존에 속 형질로 언급되지 않았던 우상 또는 세립상의 돌기를 갖는 화분괴의 표면무늬는 왜박주가리속 전체를 특징짓는 형질로 가치를 지니는 것으로 여겨진다. 한편, 기존에 속 이상의 계급에서 식별형질로 여겨지던 화분괴에 붙는 화분괴병의 위치는 나도은조롱속( Marsdenia)의 나도은조롱에서만 화분괴의 기부에 붙고( Fig. 2L), 나머지 속내에서는 상부, 아상부 또는 중부 등 화분괴의 다양한 위치에 화분괴병이 붙는 것으로 나타났다( Fig. 2A- N). 따라서 국내에 분포하는 박주가리아과에서는 본 형질이 나도은조롱속을 제외하고는 속간 한계 설정에 유용하지 않은 것으로 보인다. 한편, 덩굴박주가리( C. nipponicum Matsum. var. nipponicum)와 흑박주가리( C. nipponicum var. glabrum (Nakai) H. Hara)의 2분류군에서만 화분괴병이 화분괴의 중부에 붙는 것으로 나타났는데( Fig. 2G, H), 두 분류군은 외부형태가 유사하여 흑박주가리를 덩굴박주가리의 변종으로 취급하고 있으며, 화분괴병의 위치 외에도 화분괴, 화분괴병, 구상체의 크기 및 형태 등에서 매우 유사한 형태를 지니는 것으로 확인되었다. 구상체는 화분괴병과 더불어 직접적으로 생식에 관여하지는 않는 부위로서, 주로 수분매개체에 의해 화분 또는 화분괴를 이동시켜 주는 역할을 한다. Sugiura and Yamazaki (2005)는 구상체의 크기는 수분 시기에 따라 그 크기와 형태가 변할 수 있다고 언급하기도 했으나, 본 연구결과에서는 구상체의 형태와 크기가 분류군 내에서 다소 변이는 있으나 분류군간의 구분이 가능한 것으로 파악되었다. 구상체의 가장자리 형태는 왜박주가리속의 왜박주가리에서만 구상체의 가장자리가 결각상인 것을 본 연구에서 처음 확인하였으며( Fig. 2N), 나머지 분류군은 모두 구상체의 가장자리가 매끈한 것으로 나타났다( Fig. 2A- N). 이와 같은 특징이 왜박주가리에서만 나타나는 형질인지, 혹은 속의 식별형질인지에 대하여서는 추후 왜박주가리속의 더 많은 종의 비교 자료가 필요할 것으로 판단된다.
Chang (1986)은 한국산 박주가리아과내 박주가리속( Metaplexis)의 박주가리( M. japonica (Thunb.)와 백미꽃속의 민백미꽃( C. acuminatifolium (Decne.) Hemsl.)의 2속 2종에 대한 화분을 기재 및 측정한 바 있다. 박주가리의 경우, Chang (1986)은 화분괴의 크기를 길이 570−580 μm, 너비 250−260 μm로 기록하여, 본 연구와 거의 일치하는 것으로 나타났다( Table 1). 또한 일본산 박주가리의 화분괴 형태는 황갈색, 타원체형이며 크기는 550 × 300 μm, 구상체는 약 500 μm로 보고되고 있어( Tanaka et al., 2006), 본 조사 결과와 잘 일치하였다. 하지만 민백미꽃의 경우, 본 조사에서는 화분괴의 길이는 333−433 μm, 너비는 167−233 μm로 조사되었으나( Table 1), Chang (1986)은 140−250 μm, 너비를 150−160 μm로 보고하여 화분괴의 크기에서 본 연구 결과와 비교적 큰 차이를 보였다. 한편 Ikuse (1956)의 일본산 민백미꽃의 경우, 화분괴의 크기를 236 × 362 μm라고 보고하였으며, 이는 본 연구 결과의 범위 내에 있다. 따라서 크기에서 차이를 나타낸 Chang (1986)의 결과는 측정 오류로 판단된다.
화분괴, 화분괴병 및 구상체의 양적 형질만으로는 속간 구별은 어려웠다. 하지만 왜박주가리속은 화분괴이동체 전체적인 크기가 대체로 작은 것으로 나타났다( Table 1, Fig. 2N). 이는 왜박주가리속의 화분괴는 pendulous한 형태를 하며, 작고 타원체형에서 거의 구형인 형태를 가지고 화분괴병은 매우 가늘고 짧으며 구상체는 작고 약하다고 언급한 Endress and Bruyns (2000)와 Shah and Ahmad (2014)의 결과와도 일치하였다. 또한 박주가리속과 나도은조롱속은 백미꽃속에 비해 전체적인 크기가 비슷하거나 큰 것으로 나타났다( Table 1, Fig. 2). 화분괴, 화분괴병 및 구상체의 양적 형질은 분류군 간에 크기가 다양하게 나타나 종간 식별형질로서 유용한 것으로 확인되었다. 결론적으로, 한국산 박주가리아과에 속하는 4속 14분류군의 화분괴이동체의 형태적인 특징들을 SM과 SEM을 이용하여 비교 및 분석해 본 결과, 화분괴이동체의 형태학적 형질들은 다양하게 나타났으며, 그 중 화분괴의 방향 및 화분괴 표면의 세립상 무늬는 속간 식별형질로서 유용한 것으로 나타났다. 하지만 한국산 박주가리아과의 4속 중 백미꽃속을 제외한 3속에 각각 1종씩만 분포하고 있어, 화분학적 형질만으로 진화관계 또는 분류체계를 논하기는 어려운 것으로 생각된다. 추후 다양한 분류군의 화분괴이동체 조사를 통하여 좀 더 면밀한 조사가 요구된다.
ACKNOWLEDGMENTS
본 연구는 2014년 안동대학교 교내 연구비 지원에 의해 수행되었습니다.
Fig. 1.
The morphology and measurements of the pollinarium structure. a: the length of pollinium, b: the width of pollinium, c: the length of translator-arm, d: the length of corpusculum, e: the width of corpusculum (refer Table 1). 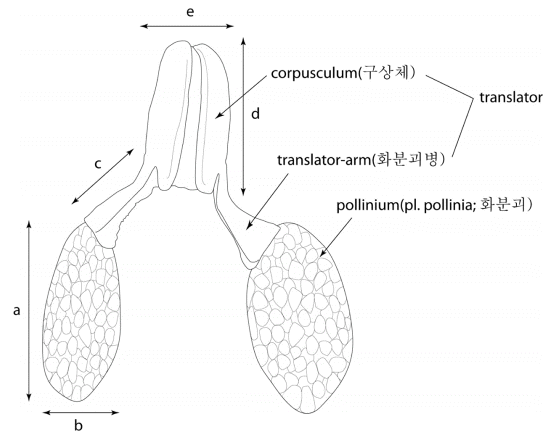
Fig. 2.
The pollinarium shapes of Korean Asclepiadoideae species observed by stereo microscope. A. Cynanchum acuminatifolium; B. C. amplexicaule; C. C. atratum; D. C. chinense; E. C. inamoenum; F. C. japonicum; G. C. nipponicum; H. C. nipponicum var. glabrum; I. C. paniculatum; J. C. volubile; K. C. wilfordii; L. Marsdenia tomentosa; M. Metaplexis japonica; N. Tylophora floribunda. 
Fig. 3.
The surface features of pollinia of Korean Asclepiadoideae species observed by SEM. A. Cynanchum acuminatifolium; B. C. amplexicaule; C. C. atratum; D. C. chinense; E. C. inamoenum; F. C. japonicum; G. C. nipponicum; H. C. nipponicum var. glabrum; I. C. paniculatum; J. C. volubile; K. C. wilfordii; L. Marsdenia tomentosa; M. Metaplexis japonica; N. Tylophora floribunda. Scale bars A−M = 10 µm, N = 5 µm. 
Table 1.
Pollinarium characters of 14 Asclepiadoideae taxa in Korea.
|
Taxon |
pollinium
|
translator-arm
|
corpusculum
|
|
*length (a) |
*width (b) |
ratio |
shape |
surface pattern |
orientation of pollinium |
*length (c) |
attachment to pollinium |
*length (d) |
*width (e) |
ratio |
shape |
type of margin |
|
Cynanchum acuminatifolium (Decne.) Hemsl. |
367 (400) 433 |
267 (276) 300 |
1.4 |
broadly ellipsoid |
psilate |
erect |
167 (187) 200 |
apical |
367 (400) 433 |
233 (260) 267 |
1.5 |
broadly ellipsoid |
entire |
|
C. amplexicaule (Siebold & Zucc.) Hemsl. |
300 (315) 333 |
83 (104) 133 |
3.0 |
narrowly ellipsoid |
psilate |
erect |
67 (96) 133 |
apical |
167 (188) 233 |
83 (99) 100 |
1.9 |
obovoid |
entire |
|
C. atratum Bunge |
333 (387) 433 |
167 (200) 233 |
1.9 |
ellipsoid |
psilate |
erect |
100 (117) 133 |
apical |
333 (407) 433 |
133 (153) 167 |
2.7 |
ellipsoid |
entire |
|
C. chinense R.Br. |
233 (283) 333 |
100 (130) 167 |
2.2 |
ellipsoid |
psilate |
erect |
67 (67) 67 |
apical |
167 (220) 300 |
100 (119) 133 |
1.8 |
ovoid |
entire |
|
C. inamoenum (Maxim.) Loes. |
133 (147) 167 |
67 (67) 67 |
2.2 |
ellipsoid |
reticulate |
erect |
100 (107) 133 |
apical |
200 (207) 233 |
67 (73) 83 |
2.8 |
narrowly ellipsoid |
entire |
|
C. japonicum C. Morren & Decne. |
267 (300) 333 |
167 (180) 200 |
1.7 |
ellipsoid |
reticulate |
erect |
67 (77) 83 |
apical |
233 (240) 267 |
100 (107) 133 |
2.2 |
narrowly ellipsoid |
entire |
|
C. nipponicum Matsum. |
300 (313) 333 |
167 (193) 233 |
1.6 |
broadly ellipsoid |
sub-reticulate |
erect |
100 (113) 133 |
middle |
333 (340) 367 |
167 (173) 200 |
2.0 |
broadly ovoid |
entire |
|
C. nipponicum var. glabrum (Nakai) H. Hara |
267 (300) 333 |
167 (183) 200 |
1.6 |
broadly ellipsoid |
sub-reticulate |
erect |
100 (123) 133 |
middle |
333 (343) 367 |
200 (207) 233 |
1.7 |
broadly ovoid |
entire |
|
C. paniculatum (Bunge) Kitag. |
367 (393) 433 |
167 (173) 200 |
2.3 |
ellipsoid |
reticulate |
erect |
100 (127) 133 |
apical |
233 (240) 267 |
117 (130) 133 |
1.8 |
ellipsoid |
entire |
|
C. volubile (Maxim.) Hemsl. |
200 (238) 267 |
100 (135) 167 |
1.8 |
ellipsoid |
sub-reticulate |
erect |
50 (66) 67 |
apical |
233 (260) 267 |
100 (138) 167 |
1.9 |
ovoid |
entire |
|
C. wilfordii (Maxim.) Hemsl. |
400 (420) 433 |
183 (203) 233 |
2.1 |
ellipsoid |
psilate |
erect |
117 (140) 167 |
apical |
300 (310) 333 |
100 (127) 133 |
2.4 |
narrowly ellipsoid |
entire |
|
Marsdenia tomentosa Morren & Decne. |
500 (547) 633 |
200 (205) 233 |
2.7 |
ellipsoid |
sub-reticulate |
erect |
133 (171) 217 |
basal |
300 (332) 367 |
167 (172) 200 |
1.9 |
obovate |
entire |
|
Metaplexis japonica (Thunb.) Makino |
467 (500) 567 |
233 (276) 300 |
1.8 |
ellipsoid |
psilate |
erect |
133 (154) 183 |
sub-apical |
400 (427) 467 |
133 (166) 200 |
2.6 |
narrowly ellipsoid |
entire |
|
Tylophora floribunda Miq. |
63 (94) 125 |
125 (145) 150 |
1.5 |
ellipsoid |
granulate |
transverse |
13 (25) 38 |
apical |
50 (66) 75 |
38 (60) 75 |
1.1 |
obovate |
incised |
Literature Cited
Bookman, SS. 1981. The floral morphology of Asclepias speciosa (Asclepiadaceae) in relation to pollination and a clarification in terminology for the genus. American Journal of Botany 68: 675-679.  Brown, R. 1810. On the Asclepiadeae, a natural order of plants separated from the Apocineae of Jussieu. Wernerian Natural History Society 1: 12-78. Chang., NK. 1986. Illustrated Flora & Fauna of Korea. Vol. 29. Pollens. the Ministry of Education, Seoul. (in Korean).
Civeyrel, L. Thomas, AL. Ferguson, K and Chase, MW. 1998. Critical reexamination of palynological characters used to delimit Asclepiadaceae in comparison to the molecular phylogeny obtained from plastid matK sequences. Molecular Phylogenetics and Evolution 9: 517-527.  Demeter, K. 1922. Vergleichende Asclepiadeenstudien. Flora 115: 130-176.
Endress, ME. 2004. Apocynaceae: Brown and now. Telopea 10: 525-541.
Endress, ME and Bruyns, PV. 2004. A revised classification of the Apocynaceae s.l. The Botanical Review 66: 1-56. Hallier, H. 1905. Provisional scheme of the natural (phylogenetic) system of flowering plants. New Phytologist 4: 151-162. Harder, LD and Johnson, SD. 2008. Function and evolution of aggregated pollen in angiosperms. International Journal of Plant Sciences 169: 59-78. Ikuse, M. 1956. Pollen Grains of Japan. Jirokawa Publishing Co, Tokyo. (in Japanese).
Johnson, SD and Edwards, T. 2000. The structure and function of orchid pollinaria. Plant Systematics and Evolution 222: 243-269. Kunze, H. 1993. Evolution of the translator in Periplocaceae and Asclepiadaceae. Plant Systematics and Evolution 185: 99-122. Lee, TB. 2003. Coloured Flora of Korea, Vol I, II Hayangmunsa; Seoul. (in Korean).
Lee, WB. 2007. Asclepiadaceae, In2016-03-29 The Genera of Vascular Plants. Park, CW (ed.), Academy Publ. Co, Seoul. 779-783.
Lee, WT. 1996. Coloured Standard Illustrations of Korean Plants Academy Publishing Co; Seoul. (in Korean).
Lee, YN. 2006. New Flora of Korea, Vol. I, II. Kyohak publishing Co., Ltd, Seoul. (in Korean).
Meve, U. 2002. Species numbers and progress in asclepiad taxonomy. Kew Bulletin 57: 459-464. Meve, U. Omlor, R and Liede, S. 2002. A new combination in Tylophora (Apocynaceae, Asclepiadoideae) from the Philippines. Systematics and Geography of Plants 72: 27-32.
Safwat, FM. 1962. The floral morphology of Secamone and the evolution of the pollinating apparatus in Asclepiadaceae. Annals of the Missouri Botanical Garden 49: 95-129. Shah, SA and Ahmad, M. 2014. Reassessing palynological characters in the subfamilies Periplocoideae and Asclepiadoideae (Apocynaceae): taxonomic and evolutionary implications. Palynology 38: 224-234. Singer, RB. Gravendeel, B. Cross, H and Ramirez, SR. 2008. The use of orchid pollinia or pollinaria for taxonomic identification. Selbyana 29: 6-19.
Sreenath, KP. Ramakrishna, TM and Babu, TP. 2012. Perspective on polinial apparatus & carriers of Asclepiadaceae sensu lato. Global Journal of Bio-Science & Biotechnology 1: 45-53.
Struwe, L. Albert, VA and Bremer, B. 1994. Cladistics and family level classification of Gentianales. Cladistics 10: 175-206. Sugiura, S and Yamazaki, K. 2005. Moth pollination of Metaplexis japonica (Apocynaceae): pollinaria transfer on the tip of the proboscis. Journal of Plant Research 118: 257-262. Swarupa, K. Mangaly, JK. Sonny, TK. Kishorekumar, K and Basha, SC. 1996. The subfamilial and tribal classification of the family Asclepiadaceae. Botanical Journal of the Linnean Society 120: 327-369. Tanaka, H. Hatano, T. Kaneko, N. Kawachino, S. Kitamura, O. Suzuki, Y. Tada, T and Yaoi, Y. 2006. Andromonoecious sex expression of flowers and pollinia delivery by insects in a Japanese milkweed Metaplexis japonica (Asclepiadaceae), with special reference to its floral morphology. Plant Species Biology 21: 193-199. Tseng, TH. Chao, ST and Lin, HW. 2011. A new species of Tylophora from coral reef areas in Hengchun Peninsula, Taiwan, China. Journal of Systematics and Evolution 49: 162. Thorne, RF. 1976. A phylogenetic classification of the Angiospermae. Evolutionary Biology 9: 35-106. Thorne, RF. 1992. An updated phylogenetic classification of the flowering plants. Aliso 13: 365-389.
Verhoeven, RL. Liede, S and Endress, ME. 2003. The tribal position of Fockea and Cibirhiza (Apocynaceae: Asclepiadoideae): evidence from pollinium structure and cpDNA sequence data. Grana 42: 70-81.
Wyatt, R. 1978. Experimental evidence concerning the role of the corpusculum in Asclepias pollination. Systematic Botany 3: 313-321.
|
|











