배추과(Brassicaceae) 식물은 전 세계적으로 338속 3,700여종이 분포하며(Al-Shehbaz et al., 2006a), 주로 온대지역에 생육하는 것으로 알려져 있다(Zhou et al., 2001). 이 중 황새냉이속(Cardamine L.)은 아시아, 북아메리카, 유럽을 중심으로 200여종이 보고되어 있으며(Zhou et al., 2001; Warwick, 2006), 히말라야 지역과 더불어 동북아시아 지역이 종분화의 중심지로 인식되고 있다(Al-Shehbaz, 1988; Lihova et al., 2006). 동북아시아 지역에는 극동러시아 13종(Komarov, 1970), 몽고 7종(Zhou et al., 2001), 중국 50종(Zhou et al., 2001; Chen et al., 2008), 대만 5종(Ying, 1996), 일본에는 17종이 분포하는 것으로 알려져 있으며(Al-Shehbaz et al., 2006b), 한국에는 가장 최근 연구에서 15종이 자생하는 것으로 기록하고 있다(Song, 2011).
애기황새냉이는 최초, 중국 길림성 지역에서 채집된 표본(Fig. 1-A)에 기초하여 Cardamine parviflora var. manshurica Kom. (Komarov, 1903)로 발표되었으나 다수의 학자들에 의해 Cardamine parviflora L.의 synonym으로 처리되고 있다(Im, 1997; Zhou et al., 2001; Al-Shehbaz et al., 2006b; Warwick et al., 2006; Oh, 2007). 그러나 Kudoh et al. (2006)은 일본을 포함한 동북아시아 지역에서 C. parviflora로 동정된 식물은 유럽 지역의 것과 형태적으로 상이하다고 주장하면서, 그 분류군이 황새냉이(C. flexuosa With.)와 매우 유사하지만 꽃과 열매의 크기가 작고 개화기가 빠른 특징을 가진다고 기록하였다. 한편, Lee (1996)는 본 분류군을 별도의 종으로 인정하여 C. manshurica (Kom) Nakai를 정명으로 채택하고 ‘애기황새냉이’라고 신칭하였으나, 현재까지 본 분류군은 황새냉이(C. flexuosa), 좁쌀냉이(C. fallax (O.E.Schulz) Nakai), 큰황새냉이(C. scutata Thunb.) 등으로 오동정되고 있다.
따라서, 본 연구에서는 기준표본과 원기재문을 바탕으로 애기황새냉이의 명확한 실체를 파악하고, 전반적인 형태형질의 고찰을 통해 변이의 한계를 규명하며, 분자유전학적 연구를 통해 본 분류군의 분류학적 위치를 재검토하고자 한다.
재료 및 방법
문헌 및 표본조사
애기황새냉이의 명확한 실체를 파악하기 위해 기본종인 C. parviflora var. manshurica의 기준표본을 조사한 후, 원기재문을 비롯한 국내炤‹ 도감, 식물지, 논문 등의 문헌의 내용을 검토하였다. 또한 국내 및 해외표본관에 소장된 황새냉이 속 표본들을 재동정하여 이용하였다. 관찰표본의 정보는 별첨하였으며, 해당표본관의 약자는 Index herbarium (http://sweetgum.nybg.org/ih/)을 기준으로 작성하였다(Appendix).
외부형태학적 연구
각 지역에서 채집된 표본 및 생체(Table 1) 등을 이용하여 각 분류군들의 형태형질을 관찰하였다. 거시적인 형질(Macro characters)은 육안 및 디지털카메라(Body: Nikon D7000, Japan; Lens: Nikon 105 mm f/2.8 AF, Japan)를 이용하여 관찰, 측정하였다. 미세형질(Micro characters)은 70% Ethanol에 고정된 개체의 특정 부위를 적출하여 1.5% glutaraldehyde에 전처리 후, 0.1M phosphate buffer (pH 6.8)로 2회 세척, 30−100% ethanol로 단계적인 탈수과정을 거쳐 isoamylacetate로 치환하였다. 이를 Ciritical point dryer (HCP-2)로 건조한 후, ion sputter로 gold coating(thickness: 200−250Å)하여 주사전자현미경(Scanning electron microscope: Jeol JSM-6300, Japan, 15−20 Kv, working distance: 15−25 mm)으로 관찰 및 촬영하였다. 또한 2007년부터 2014년까지 생육지(Table 1) 관찰을 통해 변이를 파악하였다. 연구결과는 기재문에 상세히 기록하였으며, 근연분류군과의 비교를 통하여 검색표를 작성하였다.
분자유전학적 연구
근연분류군과의 분자유전학적 계통을 연구하기 위해 nrDNA의 ITS region을 비교 분석하였다. 연구에 사용된 시료는 한국과 일본 등지에서 채집한 생체 또는 표본을 이용하였다(Table 2).
DNA extraction: 약 100mg 신선한 조직 또는 약 20mg의 건조된 잎을 조직파쇄기(Tissuelyser , Qiagen, Germany)를 이용하여 잘게 파쇄한 후, Plant DNA Extraction Kit (Cat. No. YGP100:Real Biotech Co., Taiwan)를 이용하여 Total genomic DNA를 추출하였다. 추출된 DNA는 1% agarose gel상에서 전기영동 후, EtBr 대체 시약(Loadingstar, Dyin Bio, Korea)으로 염색하여 UV 조명하에서 size maker와 상대 비교를 통하여 Length 및 Quality를 확인하였고, 흡광도를 측정(NanoDrop 2000: Thermo Fisher Scientific Inc., USA)하여 농도를 확인하였다.
PCR: PCR (Polymerase chain reaction) 반응용액의 조성은 5 ng의 주형 DNA 4 μL, 10X DiaStarTM Tag DNA buffer 5 μL, 10 mM (2.5 mM/each)의 dNTP mix 5 μL, 10 pmol의 프라이머를 각각 5 μL, seqence 전용 Tag DNA Polymerase (i-Pfu, 2.5 U/μL)를 0.5 μL에 전체 부피가 50 μL가 되도록 증류수 25.5 μL를 첨가하였다. 증폭에 이용한 프라이머는 nrDNA (nuclear ribosomal DNA)의 ITS1 (5'→3'; TCCGTAGGTGAACCTGCGG), ITS4 (5'→3'; TCCTCCGCTTATTGATATGC), ITS5 (5'→3'; GGAAGGAGAAGTCGTAACAAGG)를 이용하여 (White et al., 1990; Möler and Cronk, 1997) ITS region (ITS 1, 5.8 S, ITS 2) 구간을 증폭하였고 각각의 sequence를 비교 분석하였다. 분석된 nrDNA의 염기서열은 NCBI (http://www.ncbi.nlm.nih.gov/)에 등록하였으며(Tabel 2), 추가적으로 NCBI에 등록되어 있는 근연종 및 군외군(Outgroup)의 sequence (Table 3)를 포함하여 Neighbor-Joining 및 Maximum Likelihood 계통수를 작성하였다(CLC Main Workbench Version 8.0, Qiagen, U.S.).
결과 및 고찰
문헌 및 표본 조사
1) Cardamine parviflora var. manshurica Kom.의 실체
애기황새냉이의 기본명인 C. parviflora var. manshurica Kom. (Komarov, 1903)는 중국 길림성 지역에서 채집된 표본에 기초하여 신변종으로 발표되었으며 기준표본은 LE(V. L. Komarov Botanical Institute)와 P (Museum National d’Histoire Naturelle)에 소장되어 있다. 국제명명규약(ICN; International Code of Nomenclature for algae, fungi, and plants (Melbourne Code))의 Art 40, Note 1 (2011, http://www.iapt-taxon.org/nomen/main.php)에 의거 동가기준표본(syntypes)으로 확인되었으며(McNeill, 2014), 이중 P에 소장된 표본을 확인하였다(Fig. 1-A). 원기재문에 따르면 ‘높이 8−14 cm, 잎은 3−5장의 소엽을 가지고 0−3개의 열편을 가진 도란형, 꽃은 3−6개, 화경은 약 2 mm, 장각과는 길이 12 mm, 폭 0.5−1mm가량, 암술대가 짧으며, 둔두’인 특징을 가지는 것으로 기록되어 있으며, 이 특징은 기준표본에서도 뚜렷하게 나타난다. Marhold (1995)에 따르면 C. parviflora는 소엽이 3−11 쌍이고 소엽병이 없으며 전연인 특징이 나타나는 반면, C. hirsuta는 로제트상의 기부엽이 발달하고 소엽은 3−10 쌍이며 수술이 거의 대부분 4개이고 과병은 화축에 붙는 특징을 갖는다고 하였다. 또한 C. flexuosa는 로제트상의 기부엽이 발달하고 소엽은 10쌍 이하라고 보고하였다. 그러나 C. parviflora로 처리되고 있는 C. parviflora var. manshurica는 로제트상의 기부엽이 발달하지 않고 기부의 소엽이 0−3쌍으로, 소엽병이 뚜렷하며 엽신에는 열편이 있다. 수술은 6개이며 과병이 완전히 벌어져 있다(Table 4).
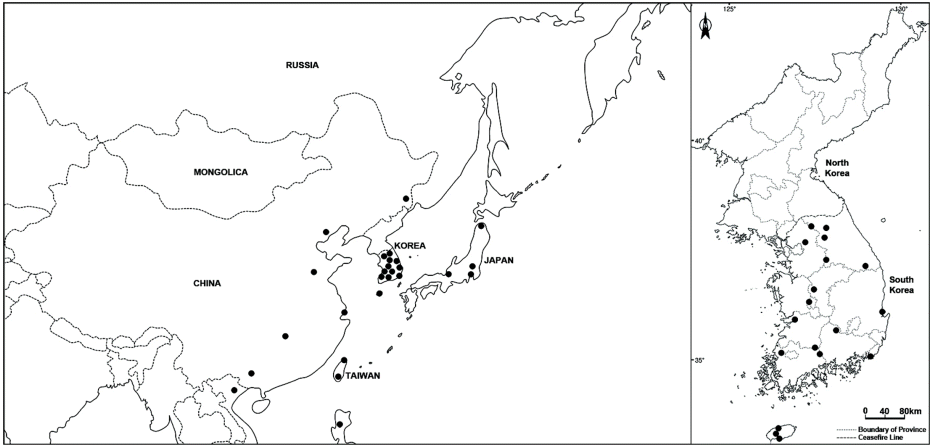
본 분류군은 국내에서 비교적 흔하게 자생하는 것으로 확인되었으며, 국내표본관에는 황새냉이(C. flexuosa), 좁쌀냉이(C. fallax), 큰황새냉이(C. scutata) 등으로, 해외표본관에는 C. flexuosa, C. parviflora, C. hirsuta 등으로 오동정된 채 소장되어 있었다. Komarov (1903)는 중국, 일본, 러시아(Altai, Kavkaz)를 비롯하여 유럽 남부까지 분포한다고 기록하였으나, 표본조사결과 러시아와 유럽지역의 표본은 확인하지 못하였다(Fig. 2, Appendix).
2) Cardamine manshurica (Kom.) Nakai의 정당성
Nakai는 1909년 6월 4일, 부산 용두산에서 채집한 C. parviflora var. manshurica의 표본을 ‘C. flexuosa var. manshurica Kom.’로 동정하여 기록하였으며, Flora Koreana II에 C. parviflora var. manshurica를 C. flexuosa var. manshurica(Kom.)의 이명(synonym)으로 처리하였다(Nakai, 1911). Tyosen Syokubutsu (Nakai, 1914)에는 ‘Cardamine manshurica Nakai’라는 학명이 기록되어 있으며, 간략한 도해와 일본어로 된 기재문이 수록되어 있다(Fig. 1-B). 도해는 Komarov에 의해 채집된 C. parviflora var. manshurica의 기준표본에서 드러나는 특징과 원기재문의 내용(3−5장의 소엽, 0−3개의 열편을 가진 도란형, 꽃이 3−6개)이 비교적 잘 묘사되어 있으며 일본어로 기술되어 있는 기재문의 내용(줄기는 짧고 작으며, 길이는 4−5촌을 넘지 않고, 잎 또한 작다. 황새냉이의 일종이나 편의상 구분)도 마찬가지로 부합된다. ICN의 Art 39에 의하면, 1935년 이전에는 라틴어 이외의 다른 언어로 기재되어 있을지라도 합법명으로 인정된다. 본 학명은 The International Plant Names Index (http://www.ipni.org), Tropicos (http://www.tropicos.org), The Plant List (http://www.theplantlist.org)에는 ‘Cardamine mans-hurica (Kom.) Nakai’로 기록되어 있으며, 조선식물지경계 (Nakai, 1952)에서도 ‘Cardamine manshurica (Komarov) Nakai’로 기록하고 있다. 그러나 Nakai가 직접적으로 기본명을 명시한 문헌은 찾을 수 없었기 때문에, Komarov (1903)가 발표한 C. parviflora var. manshurica Kom.와 C. flexuosa var. manshurica Kom. 중 어느 것이 basionym인지 명확하지 않다. 그러나 국제명명규약 Art 41.4항에 준하여 1953년 이전에 공표된 학명에 대해서는 기본명에 대한 명확한 인용이 없더라도 신조합된 이름으로 처리될 수 있으며, 후학들의 견해에 따라 C. parviflora var. manshurica Kom.을 기본명으로 간주할 수 있다(Kanchi N. Gandhi, personal communication).
3) 국명에 관한 논의
Nakai (1914)는 C. manshurica (Kom.) Nakai에 작고 귀여운 황새냉이라는 뜻을 가진 일본명 ‘ひめたねつけばな’을 신칭하였다. Lee (1976)는 Nakai가 1909년 부산 용두산에서 채집한 표본의 지명을 따서 ‘용머리냉이’라고 하였으며, Lee (1996)는 일본어를 번역한 ‘애기황새냉이’로 신칭하였다. 근연분류군보다 일반적으로 키가 작고 꽃이 소형인 형태적 특징을 고려할 때, 애기황새냉이를 국명으로 사용하는 것이 바람직하다고 판단된다. 한편, 유럽에 분포하는 C. parviflora의 국명으로 사용되어 왔던 ‘좀냉이’(Koh, 1993; Korea national arboretum and the plant taxonomic society of Korea, 2007)와 ‘말황새냉이’(Im, 1997; Oh, 2007)는 사용하지 않는 것이 옳다고 판단된다.
외부형태학적 연구
Kudoh et al. (2006)이 언급한 바와 같이 본 분류군은 외부형태학적으로 황새냉이와 가장 유사하지만, 화조(花爪, claw)에 거치가 없고 중앙밀선이 거의 퇴화되어 있으며 종자의 선단부가 얇아지지 않는다(Fig. 3). 특히 한국의 중부 고산지대에서는 주로 5월에 개화하기 시작하여 10월경까지, 남부 저지대에서는 2월부터 12월까지 개화와 결실을 지속적으로 반복하여 1년 동안 3−5회 세대교번이 이루어지는 반면, 황새냉이를 비롯한 근연분류군은 연 1회, 드물게 2회 개화하는 것으로 확인되었다. 상기 특징들에 의해 본 분류군은 근연종과 종 수준에서 명확하게 구분되었다. 한편, 본 연구를 통해 관찰된 세부형질들은 기재문에 상세히 기록하였다.
분자생물학적 연구
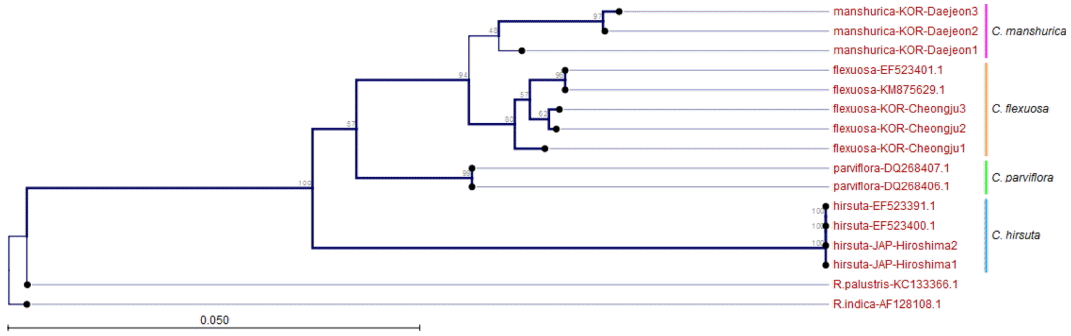
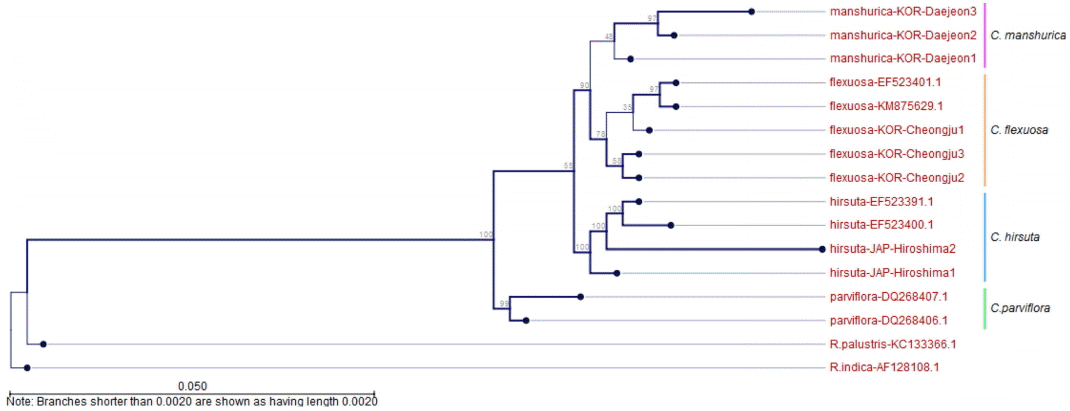
분석된 애기황새냉이의 nrDNA 염기서열 전체길이는 683 bp이었으며, 배추과(Brassicaceae)의 모델식물인 애기장대를 기준으로 정렬하여 비교하였다. 정렬된 구간별 길이는 ITS 1이 270 bp, 5.8S가 164 bp, ITS 2는 215 bp로 ITS 1 구간이 ITS 2 구간에 비해 긴 것으로 나타났다. 기존에 통합되어 왔던 C. parviflora와 근연종으로 알려졌던 C. hirsuta, 외부형태적으로 가장 근연한 것으로 판단되는 C. flexuosa를 내군(Ingroup)으로 설정하였고, Lihová et al. (2006), Ali et al. (2010), Song (2011) 등과 같이 개갓냉이속(Genus Rorippa) 식물을 외군(Outgroup)으로 설정하였다. Neighbor-Joining (NJ) Tree에서는 일본과 유럽의 C. hirsuta는 100%의 bootstrap value로 유집되어 C. manshurica와 유연관계가 가장 먼 것으로 나타났고, 유럽에 분포하는 C. parviflora는 C. flxuosa 및 C. manshurica 그룹과 57%의 bootstrap value로 유집되었다. C. flxuosa와 C. manshurica는 94%의 bootstrap value로 별도의 분계조를 형성하였다(Fig. 4). Maximum Likelihood (ML) Tree에서도 bootstrap value의 차이는 다소 있었으나 동일한 분계조를 형성하는 것으로 확인되었다(Fig. 5). 따라서 분자생물학적 결과는 본 종이 유럽에 분포하는 C. parviflora 의 종하분류군 또는 이명(synonym)으로 처리될 수 없음을 지지하였고, C. hirsuta와는 유연관계가 먼 것으로 확인되었으며 C. flexuosa와 근연종임을 알 수 있었다. 한편, C. flexuosa와 C. manshurica는 동일한 집단 내에서도 개체에 따른 sequence의 변이가 다양하게 나타나며, 이러한 이유를 Lihová et al. (2006), Song(2011)은 C. flexuosa complex에서 빈번하게 이루어지는 잡종화 현상과 배수화 현상에 기인한다고 하였다. 그러나 C. manshurica의 세대교번의 횟수, 동일 지역에서의 개화시기 차이, 분포역에 따른 환경의 차이 등을 종합적으로 고려하였을 때, 집단 및 개체에 따라 유전자 변이가 일어날 확률이 다른 분류군에 비해 월등하게 높기 때문이라고 판단된다.
향후 기준채집지인 만주 지역의 표본과 종자를 확보하여 외부형태학적연구 및 분자생물학적연구를 추가적으로 수행하여 비교한다면 애기황새냉이를 둘러싼 분류학적 논란을 보다 확실하게 정리할 수 있을 것으로 기대할 수 있다.
분류학적 처리
C. parviflora var. manshurica Kom., Trudy Imp. S.-Petersbursg Bot. Sada 22: 371. 1903. Type: China. Provincia Kirin, Badao-chezsa fluv. Suifun (Prov. Jilin, Suifen river), 20 Jun. 1896, V. L. Komarov s.n. (Syntypes: LE, not seen; P, Photo!).
Herbs, annual or biannual, 2−30 cm tall, nearly glabrous or hirsute. Taproots slender or rarely slightly fleshy, mostly with fibrous adventitious roots. Stems usually branched basally to apically, erect to decumbent, leafy, often purple at base, nearly glabrous to downward hirsute. Leaves only cauline. Cauline leaves mostly compound or rarely simple in basalmost ones in annual plants, 1.3−5.5 cm, glabrous or hirsute; petiole 0.3−4 cm, ridged; terminal leaflet petiolulate, obovate to reniform, usually 3−5-lobate, 0.5−3.3 × 0.4−1.4 cm, cordate at base, irregularly crenate or entire, ciliate or not in margin, rounded or obtuse in apex; lateral leaflets 1−3 pairs, petiolule 0.1−0.5 cm, similar to terminal one in shape, 0.3−2.5 × 0.1−1.1 cm. Inflorescences raceme, few to many-flowered; pedicels green to purple, stipular glands absent, glabrous. Flowers symmetrical; sepals 4, green to purple, lanceolate, concave, 1.5−2.3 × 0.5−0.7 mm, almost glabrous or rarely sparsely hirsute abaxially, petals 4, white, spatulate to oblanceolate, 2.0−3.0 × 0.8−1.2 mm, apex rounded; basal claw attenuate, entire in margin; stamens 6, tetradynamous, superior ones 2−2.7 mm, inferior ones 1.5−2.2 mm; filaments white, ascending, linear; anthers yellow, elliptical to cordate, 0.4−0.5 × 0.5−0.6 mm; nectar glands 4, rarely 2 or 6, separate, median glands 2 or rarely degenerated, linguiform or rarely papilla shape, less than 0.1 × 0.1 mm, lateral glands 2 or rarely 4, linguiform, less than 0.1 × 0.1 mm; ovary green, 1.8−2.3 × 0.5 mm, glabrous, midrib smooth; ovules 14−20 per locule; style obsolete, ca. 0.15 mm; stigma capitate, ca. 0.8 mm diam. Siliques green to purple, 8.0−23.0 × 1.3−1.5 mm, ca. 1 mm in thickness; stalks erect or divaricate, straight to ascenidng, deviated or not toward shady place; valves glabrous or pubescent, coiled or not at dehiscence; beak condensed, 0.5−0.7 mm. Seeds continuous, yellowish to greenish brown, elliptical, oblong, oval or ovate, 0.9−1.3 × 0.7−0.9 × 0.3−0.5 mm, smooth in margin; seed coat reticulate, anticlinal cell walls straight, inter cellular channels convex.
Distribution and habitat: China, Japan, Taiwan, Philippines, Vietnam and Korea (All provinces). Wet places around valleys and streams, riversides, farmlands, roadsides and bare grounds.
Phenology: Flowering from January to December.
종 검색표
1. 소엽병이 없고, 정소엽은 일반적으로 전연이며, 선형 또는 도피침형, 드물게 피침상 도란형이다 ·· C. parviflora
1. 소엽병은 약 1−2 mm, 정소엽은 흔히 3−7개로 갈라지며, 신장형 또는 도란형이다.
2. 수술은 일반적으로 4개이다. 과병은 화축에 붙어 있고, 과피의 선단부가 원두이다 ···················· C. hirsuta
2. 수술은 6개이다. 과병은 화축에서 떨어져 있고, 과피의 선단부가 예두이다.
3. 기부엽은 로제트상이고, 중앙밀선은 길이와 폭이 0.1 mm 이상이다. 화조는 기부에서 급격하게 좁아 지고 거치가 뚜렷하다 ······························ C. flexuosa
3. 기부엽은 로제트상을 이루지 않고, 중앙밀선은 0.1 mm 이하이거나 없다. 화조는 밋밋하게 좁아지고 거치가 없다 ····································· C. manshurica