Approximately 2,000 islands are found in a narrow band along the southern coastal region of Korean Peninsula including Jeju Island (Yang, 2013). This edge zone is disjunctively located on the northwestern margin of an equivalent climatic area of Japan, and is covered by various evergreen broad-leaved tree species (Satake et al., 1989; Lee and Choi, 2010). Thus, Korean populations of a given species may have an important evolutionary property (J.-H. Lee et al., 2013; Lee et al., 2014) that has guaranteed their long-term survival in East Asia (Frankham, 2005; Ouborg et al., 2006; Willi et al., 2006).
Daphne kiusiana Miq. var. kiusiana is an evergreen broad-leaved shrub endemic to Korea (mostly on Jeju Island) and Japan. However, plants on Jeju Island are sometimes regarded as a different species with D. kiusiana in Japan because of several unique morphological traits, e.g., a glabrous calyx and lobes, and elliptical leaves (c.f., D. jejudoensis M. Kim) (J.
Lee et al., 2013). Therefore, a clearer boundary of taxonomic delimitation is needed. Moreover, natural populations in Korea are threatened by anthropogenic pressures, i.e., disturbances that have caused habitat destruction due to urbanization as well as over-collecting of specimens for commercial purposes (Ro et al., 2010). For example, because of its graceful shape (resembling a bride's bouquet) and pleasant fragrance, it is commonly used as an ornamental (Ro et al., 2010). Populations there are now extremely restricted to several islands, including Geoje and Jeju, where plants are being managed as endangered organisms (Korea National Arboretum, 2008). Despite these protective measures, human activity continues to threaten their existence. Therefore, we developed a set of polymorphic microsatellite markers from D. kiusiana to resolve this debate about the taxonomic entity of Jeju populations and establish a suitable conservation strategy for Korean populations based on genetic diversity and structure.
Materials and Methods
To acquire our microsatellite library, we used a DNeasy Plant Mini Kit (Qiagen, Seoul, Korea) to extract genomic DNA from a fresh leaf of a single plant sampled at Sinpyeonggotjawal, Jeju Island, Korea (Table 1). Measurements were made with a NanoDrop ND-1000 (NanoDrop Technologies, Wilmington, DE, USA). Applying a Next Generation Sequencing (NGS) approach, we sequenced high-quality DNA (concentration: 46 ng/µL; A260/280 = 1.73; A260/230 = 1.64) using the Illumina MiSeq platform (LAS, Seoul, Korea). In all, 4,936,656 reads (2 × 300 bp) were produced by Illumina paired-end sequencing and then trimmed by Trimmomatic 0.32 (Bolger et al., 2014).
To identify the microsatellites from those reads, we screened them with SSR pipeline v. 0.951 (Miller et al., 2013). The parameters were set for detection of di-, tri-, or tetra-nucleotide motifs with flanking regions larger than 50 bp and having at least 10, 6, or 4 repeats, respectively. From this screening, we acquired 28,495 sequences that were then re-filtered in an attempt to reference-map all of the reads to each microsatellite that contained a singleton. This was accomplished with Geneious R7.1.8 (Biomatters; available from http://www.geneious.com). After discarding putative multi-copy loci with exceptionally high coverage, we selected fragments with unique patterns that had two separate alleles, few variations at the site to which a primer was attached, and no additional single nucleotide polymorphisms in the flanking region. Finally, we designed 140 primer pairs with Primer3 in Geneious R7, and synthesized with a taq sequence (5′-CACGACGTTGTAA AACGAC-3′) to enable incorporation with M13 primer labeled at the 5'-end with a 6-FAM fluorescent dye.
To test the effectiveness of the developed microsatellite loci, we collected 80 individuals of D. kiusiana from three natural populations in Korea and Japan (Table 1). To minimize the damage to this study species, only one plant per population was taken to prepare a voucher specimen that was then deposited in the herbarium at the Korea Institute of Oriental Medicine (KIOM). The DNA was extracted as described above, and PCR was conducted with a GeneAmp® PCR System 2700 Thermal Cycler (Applied Biosystems, Foster City, CA, USA), using a final volume of 10 μL that comprised 15 to 20 ng of extracted DNA, 5 µL of SolgTM 2× Taq PCR Smart-Mix I (Solgent, Daejeon, Korea), 0.01 µM forward primer, 0.2 µM reverse primer, and 0.1 µM of the M13 primer (fluorescently labeled). The PCR amplifications were performed under the following conditions: initial denaturation at 95 oC for 2 min; then 30 cycles of denaturing at 94 oC for 30 s, annealing at 53 to 58 oC for 45 s, and extension at 72 oC for 45 s; with a final extension at 72 oC for 7 min. Without dilution of fluorescently labeled PCR products, 1 µL was analyzed concurrently with the GeneScanTM-500LIZTM Size Standard (Applied Biosystems) on an ABI 3730XL sequencer (Applied Biosystems). Allele sizes were manually determined three times with GENEMAPPER 3.7 software (Applied Biosystems). The number of alleles plus values for HE (expected heterozygosity) and HO (observed heterozygosity) were obtained with GenAlEx 6 (Peakall and Smouse, 2006). Deviations from Hardy–Weinberg equilibrium (HWE) was estimated with GENEPOP 4.0 (Rousset, 2008).
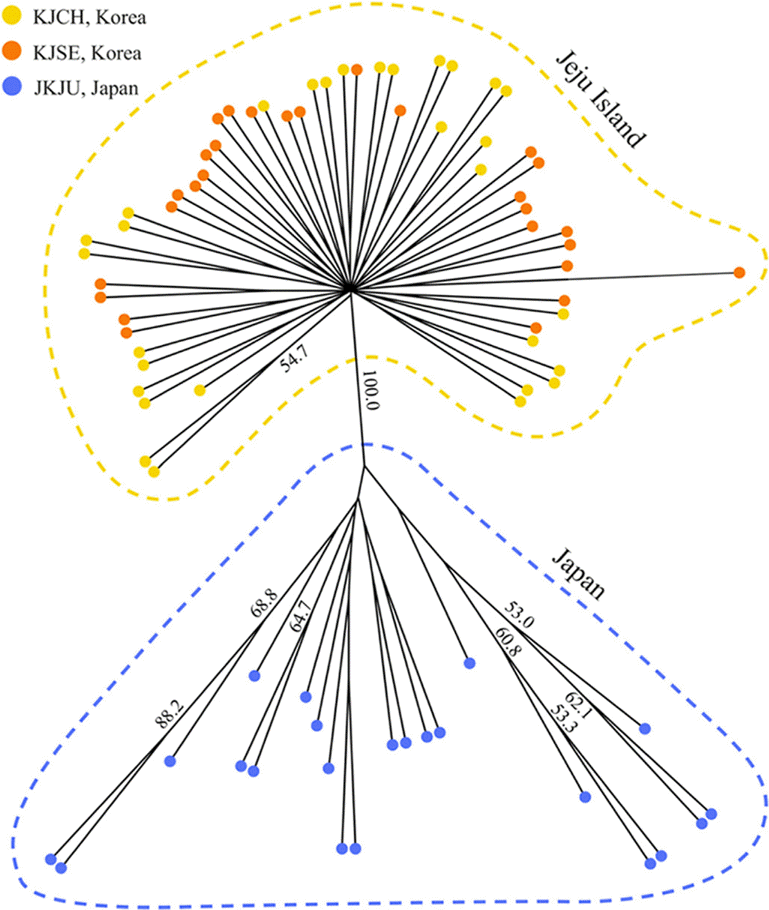
To determine the genetic association among the three populations, we quantified their relationship using a proportion of shared alleles (Dps) based on 19 loci. Only DKi134 and DKi140 were excluded. The pair-wise genetic distance (Dps) was generated with MSA software (Dieringer and Schlöterer, 2003) using a bootstrap analysis of 1,000 replicates. From these distance matrices, we constructed 50% consensus trees by the Neighbor-Joining (NJ) method, as implemented in PHYLIP ver. 3.68 (Felsenstein, 2004).
Results and Discussion
Of the developed 140 primer pairs, 21 proved to be polymorphic while the rest either were insufficiently amplified
or else produced inconsequential peaks (Table 2). Overall, the alleles numbered 2 to 7 (average of 3.90). Values for HE and HO ranged from 0.000 to 0.601 and from 0.000 to 0.700, respectively (Table 3). Among the 21 loci, two and nine were monomorphic in Jeju population KJSE and Japanese population JKJU, respectively. Loci DKi134 and DKi140 were not amplified in the Japanese population. Furthermore, nine polymorphic and monomorphic loci clearly did not overlap between Jeju Island and Japan within the size range of those alleles (Table 3). Although eight loci showed significant deviations from Hardy–Weinberg equilibrium, this was not consistent across populations. The NJ tree classified the three populations into two groups according to geographical or morphological traits (Jeju Island vs. Japan) with a remarkably high bootstrap value of 100 (Fig. 1). The microsatellite markers described here will be a powerful genetics tool for elucidating the taxonomic entity of Jeju populations, and in the organization of conservation programs planned to reduce inbreeding via artificial crossbreeding and to minimize loss of genetic variability in Jeju populations. Also, we expect that they will also improve our understanding about historical and contemporary gene flow, not only on a fine scale within Korea but also on a larger scale across the entire range of this species.
Table 1.
Voucher and location information for Daphne kiusiana samples used in this study. One specimen per population was deposited in the herbarium at the Korea Institute of Oriental Medicine (KIOM).
Table 2.
Characteristics of 21 microsatellite loci developed in Daphne kiusiana.
| Locus | Primer sequence (5′–3′)a |
Repeat motif |
A | Tm (°C) |
Size range (bp) |
GenBank accession No. |
|---|---|---|---|---|---|---|
| DKi019 | F: CCTAATGATTTAGACCGCT | (CT)12 | 5 | 53 | 206–232 | KT932539 |
| R: TTCTGGTGAGAAACGAAC | ||||||
| DKi020 | F: GTACAGATCTCAATTGGTCT | (CT)13 | 4 | 53 | 244–252 | KT932540 |
| R: GATAGCAATCATCCATCAAG | ||||||
| DKi021 | F: CGTTTAGGGTGTAAGTCATA | (CT)16 | 7 | 53 | 267–285 | KT932541 |
| R: TACTCGATCCATCCCTATTA | ||||||
| DKi022 | F: CAGAGACAGGATTGAACAT | (GA)13 | 6 | 53 | 197–215 | KT932542 |
| R: ATTGGATACGTGTTAGGTC | ||||||
| DKi023 | F: CAGTTGAGGATGTCATATCA | (GA)13 | 6 | 53 | 279–293 | KT932543 |
| R: GCCGAATGTGTTATTTGTC | ||||||
| DKi101 | F: AGATGCTGGTTCATGTTCG | (AC)10 | 5 | 53 | 179–193 | KT932572 |
| R: GGAACACAGTTGAACGTTG | ||||||
| DKi104 | F: CCTGCGATCTGGGTAATTT | (AT)10 | 4 | 58 | 239–251 | KT932574 |
| R: AGGCCACACAACAAGAATC | ||||||
| DKi062 | F: ATCAACAATCTACTCCTGC | (ATG)8 | 4 | 53 | 247–262 | KT932553 |
| R: CACTCCTTCCAGAACTAAG | ||||||
| DKi070 | F: AGAAGATGATGCCAAGATG | (GAA)9 | 4 | 58 | 245–254 | KT932557 |
| R: GGAAGACTTAACGGCTAAT | ||||||
| DKi072 | F: AAGGTGGGAGATTCTGATA | (GAT)8 | 2 | 53 | 199–205 | KT932558 |
| R: GAGCACTAGACAATTCCTT | ||||||
| DKi073 | F: GTCTTAGGGCTAGTGATTC | (GAT)9 | 3 | 53 | 248–263 | KT932559 |
| R: GACCGTCAAGTCTTCTATT | ||||||
| DKi082 | F: TCAAGACCAATATTGCCAG | (TGA)8 | 4 | 53 | 183–201 | KT932564 |
| R: CTCCATGAGGTGATTCATT | ||||||
| DKi117 | F: ATAGTTCTTGTGCCACCAC | (ATA)6 | 2 | 53 | 263–266 | KT932579 |
| R: TAGCCAAGTCGTTGTCTTG | ||||||
| DKi119 | F: AAGCACAATCATGGAGCAT | (CTT)6 | 7 | 53 | 172–208 | KT932580 |
| R: ATCGTCAGAACTAGCCTGA | ||||||
| DKi091 | F: TGAGAGACTCAGTACTCTC | (AGAA)5 | 3 | 53 | 309–317 | KT932569 |
| R: ACTACCACTCTCTGTTTCT | ||||||
| DKi097 | F: ACTTGTCCTCAACTTTCTC | (TTAA)5 | 3 | 53 | 242–258 | KT932571 |
| R: TATTTGATGATCAGGTCGC | ||||||
| DKi128 | F: ATAAAGCGGAATGGAGTCG | (TTGA)7 | 3 | 53 | 179–187 | KT932585 |
| R: ACAGTAGTAGGTCCCACAC | ||||||
| DKi129 | F: AGTTCAGACGCTTCAACC | (CATA)6 | 4 | 58 | 296–312 | KT932586 |
| R: CCATCCACAGAGACAGATTC | ||||||
| DKi131 | F: GAAACTCGTCTCTCTTCCTC | (AGGA)5 | 2 | 53 | 199–203 | KT932588 |
| R: GACGACTACCATAGCATAGC | ||||||
| DKi134 | F: AGTGCTTTAATGGAAGGCT | (CTAT)4 | 2 | 58 | 203–223 | KT932589 |
| R: GATCACCTCCTAACCAAGC | ||||||
| DKi140 | F: ATTCACACCCTAGTCGGAT | (TGTA)4 | 2 | 53 | 215–219 | KT932590 |
| R: GAAACAAAGCTGAGCGAAG |
Table 3.
Genetic parameters of 21 microsatellite loci for separate populations of Daphne kiusiana sampled in Jeju Island (KJCH and KJSE) and Japan (JKJU).
| Locus |
KJCHa (n = 30) |
KJSEa (n = 28) |
JKJUa (n = 22) |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | HE | HO | Sr | A | HE | HO | Sr | A | HE | HO | Sr | |
| DKi019 | 2 | 0.500 | 0.467 | 216–218 | 2 | 0.436 | 0.429 | 216–218 | 3 | 0.458 | 0.136* | 206–232 |
| DKi020 | 2 | 0.095 | 0.100 | 244–248 | 2 | 0.035 | 0.036 | 248–250 | 3 | 0.241 | 0.273 | 248–252 |
| DKi021 | 5 | 0.537 | 0.600 | 275–285 | 4 | 0.198 | 0.143* | 275–285 | 2 | 0.434 | 0.000* | 267–269 |
| DKi022 | 3 | 0.493 | 0.467 | 197–213 | 3 | 0.601 | 0.643* | 197–201 | 2 | 0.351 | 0.091* | 211–215 |
| DKi023 | 3 | 0.443 | 0.300 | 287–293 | 2 | 0.459 | 0.571 | 287–291 | 3 | 0.507 | 0.091* | 279–283 |
| DKi101 | 3 | 0.259 | 0.200* | 179–183 | 2 | 0.163 | 0.107 | 179–181 | 2 | 0.351 | 0.091* | 189–193 |
| DKi104 | 3 | 0.509 | 0.433 | 245–251 | 2 | 0.226 | 0.185 | 245–249 | 1 | 0.000 | 0.000 | 239 |
| DKi062 | 2 | 0.498 | 0.467 | 259–262 | 2 | 0.219 | 0.250 | 259–262 | 2 | 0.044 | 0.045 | 247–256 |
| DKi070 | 3 | 0.513 | 0.700 | 245–254 | 2 | 0.494 | 0.464 | 245–251 | 1 | 0.000 | 0.000 | 248 |
| DKi072 | 2 | 0.444 | 0.467 | 199–205 | 2 | 0.293 | 0.286 | 199–205 | 1 | 0.000 | 0.000 | 205 |
| DKi073 | 3 | 0.496 | 0.433* | 248–263 | 2 | 0.436 | 0.357 | 248–254 | 1 | 0.000 | 0.000 | 248 |
| DKi082 | 2 | 0.064 | 0.067 | 195–198 | 3 | 0.070 | 0.071 | 195–201 | 1 | 0.000 | 0.000 | 183 |
| DKi117 | 2 | 0.339 | 0.233 | 263–266 | 2 | 0.484 | 0.464 | 263–266 | 1 | 0.000 | 0.000 | 263 |
| DKi119 | 4 | 0.585 | 0.567 | 172–208 | 4 | 0.284 | 0.321 | 178–208 | 3 | 0.433 | 0.545 | 193–199 |
| DKi091 | 2 | 0.406 | 0.433 | 313–317 | 2 | 0.357 | 0.393 | 313–317 | 1 | 0.000 | 0.000 | 309 |
| DKi097 | 2 | 0.278 | 0.133* | 242–250 | 2 | 0.316 | 0.250 | 242–250 | 1 | 0.000 | 0.000 | 258 |
| DKi128 | 3 | 0.415 | 0.367 | 179–187 | 2 | 0.497 | 0.357 | 183–187 | 2 | 0.044 | 0.045 | 179–183 |
| DKi129 | 2 | 0.206 | 0.233 | 308–312 | 1 | 0.000 | 0.000 | 308 | 2 | 0.397 | 0.545 | 296–304 |
| DKi131 | 2 | 0.480 | 0.267* | 199–203 | 2 | 0.499 | 0.464 | 199–203 | 1 | 0.000 | 0.000 | 203 |
| DKi134 | 2 | 0.444 | 0.333 | 203–223 | 2 | 0.494 | 0.607 | 203–223 | 0 | N/A | N/A | N/A |
| DKi140 | 2 | 0.095 | 0.100 | 215–219 | 1 | 0.000 | 0.000 | 219 | 0 | N/A | N/A | N/A |
A, number of alleles; HE, expected heterozygosity; HO, observed heterozygosity; Sr, size range; N/A, unavailable PCR products.
a Population symbols are the same as those used in parentheses for Table 1.