INTRODUCTION
Eleocharis R. Br. is characterized by unbranched culms, one terminal spikelet, bisexual flowers, and bristly perianths (Smith et al., 2002; Dai and Strong, 2010; Park et al., 2016). It occurs mainly in circumboreal marshes and wetlands, and includes about 250 species (González-Elizondo and Peterson, 1997; Smith et al., 2002; Dai and Strong, 2010). González-Elizondo and Peterson (1997) recognized four subgenera, seven sections, eight series, and seven subseries. In Korean Eleocharis, Oh (2007) and Park et al. (2016) recognized 13 or 15 taxa, respectively. Oh and Lee (1997) and Oh (2000) classified Korean taxa into two sections (Limnochloa (P. Beauv. & T. Lestib.) Benth. & Hook. f. and Eleocharis R. Br.) based on spikelet and achene sizes.
Eleocharis possesses holocentric chromosomes (diffuse chromosomes), which miss constricted centromeres during cell divisions (Håkansson, 1954, 1958; Hoshino, 1987; Yano et al., 2004). Chromosome numbers in the genus range broadly from 2n = 10 to 2n = 196 (x = 5), and mixoploidy is common (Hoshino, 1987; Hoshino et al., 1993; Yano et al., 2004). The sizes of chromosomes also vary in the genus from less than 0.7 μm to about 6 μm (Hoshino, 1987; Hoshino et al., 1993; Yano et al., 2004). Eleocharis taxa with small chromosome numbers generally have large chromosomes such as E. valleculosa f. setosa Ohwi (2n = 16, ca. 3–6 μm in length) whereas taxa with large chromosome numbers exhibit small chromosomes as in E. dulcis (Burm. f.) Trin. ex Hensch. (2n = ca. 196, less than 1 μm in length) (Hoshino et al., 1993; Yano et al., 2004). In addition, satellites were observed in some taxa, E. mamillata var. cycrocarpa Kitag. and E. acicularis (L.) Roem. & Schult. f. longiseta (Svenson) T. Koyama (Hoshino et al., 1993; Yano et al., 2004).
Out of 15 Korean taxa, none of them has chromosome data reported. However, a few Eleocharis taxa from Chinese and Japanese populations have been reported (Hoshino, 1987; Hoshino et al., 1993; Yano et al., 2004; Hoshino et al., 2011). E. mamillata var. cycrocarpa has chromosome numbers of 2n = 16 commonly in Chinese and Japanese populations, but two satellites are observed only in Chinese populations (Hoshino et al., 1993). This suggests chromosomal diversification within a taxon. Intraspecific diversity in chromosome number and configuration are related with diffuse centrometric chromosomes (Hoshino et al., 1993; Yano et al., 2004). Eleocharis chromosome data analyzed in a phylogenetic framework suggest that fragmentation (fission), fusion and polyploid events contributed to the species diversity in the genus (Yano et al., 2004).
In the previous investigations, several types of chromosomes have been recognized in Eleocharis. Hoshino (1987) recognized three types of chromosomes in Eleocharis: (1) round prochromosome type (bimodal variation) (E. mamillata var. cyclocarpa), (2) complex chromocenter type (gradational variation) (E. wichurae Boeckeler, E. tetraquetra Nees, E. acicularis (L.) Roem. & Schult., and E. congesta var. thermalis (Hultén) T. Koyama), and (3) sparsely diffuse type (very small, under 0.7 μm in length). Variation in chromosome numbers has also been reported in the genus and also in other genera in Cyperaceae. The chromosome variation in size and number is hypothesized in a phylogenetic framework by Yano et al. (2004). There are three types of chromosome groups supported by phylogenetic data: (1) very small chromosomes in section Limnochloa (E. dulcis, E. kuroguwai Ohwi), (2) gradient variation in size in sections Pauciflorae (Beauverd) Svenson and Eleocharis, and (3) two-sized chromosomes, bimodal karyotypes (large and small) in series Eleocharis.
We report somatic chromosome numbers of four Korean native Eleocharis taxa: E. attenuata f. laeviseta (Nakai) H. Hara, E. parvula (Roem. & Schult.) Link ex Bluff, Nees & Schauer, E. ussuriensis Zinserl. and E. valleculosa var. setosa Ohwi. Significance of their chromosome numbers is also discussed.
MATERIALS AND METHODS
Root tips were sampled in natural habitats during the field surveys in various collection sites (Table 1). Initially, root tips were pretreated in 0.002 M 8-hydroxiquinoline for over 8 h and then fixed in a mixture of ethanol and glacial acetic acid (3:1) for 12 h. After fixation, root tips were preserved in 70 % ethanol, then macerated in 1 N mol HCl for 10 min before 1 % acetic-orcein staining. Stained root tips were squared and observed at 1,000× magnification (Nikon Eclipse 50i, Nikon, Tokyo, Japan). At least three mitotic cells per individual were analyzed to confirm numbers and types of chromosomes. Drawing and photographs were made for the cells analyzed. Park et al. (2016) was used for identification, and scientific and Korean names followed those at the Korea National Institute of Biological Resources species database. Voucher specimens were saved in the Korea National Institute of Biological Resources herbarium (KB).
RESULTS AND DISCUSSION
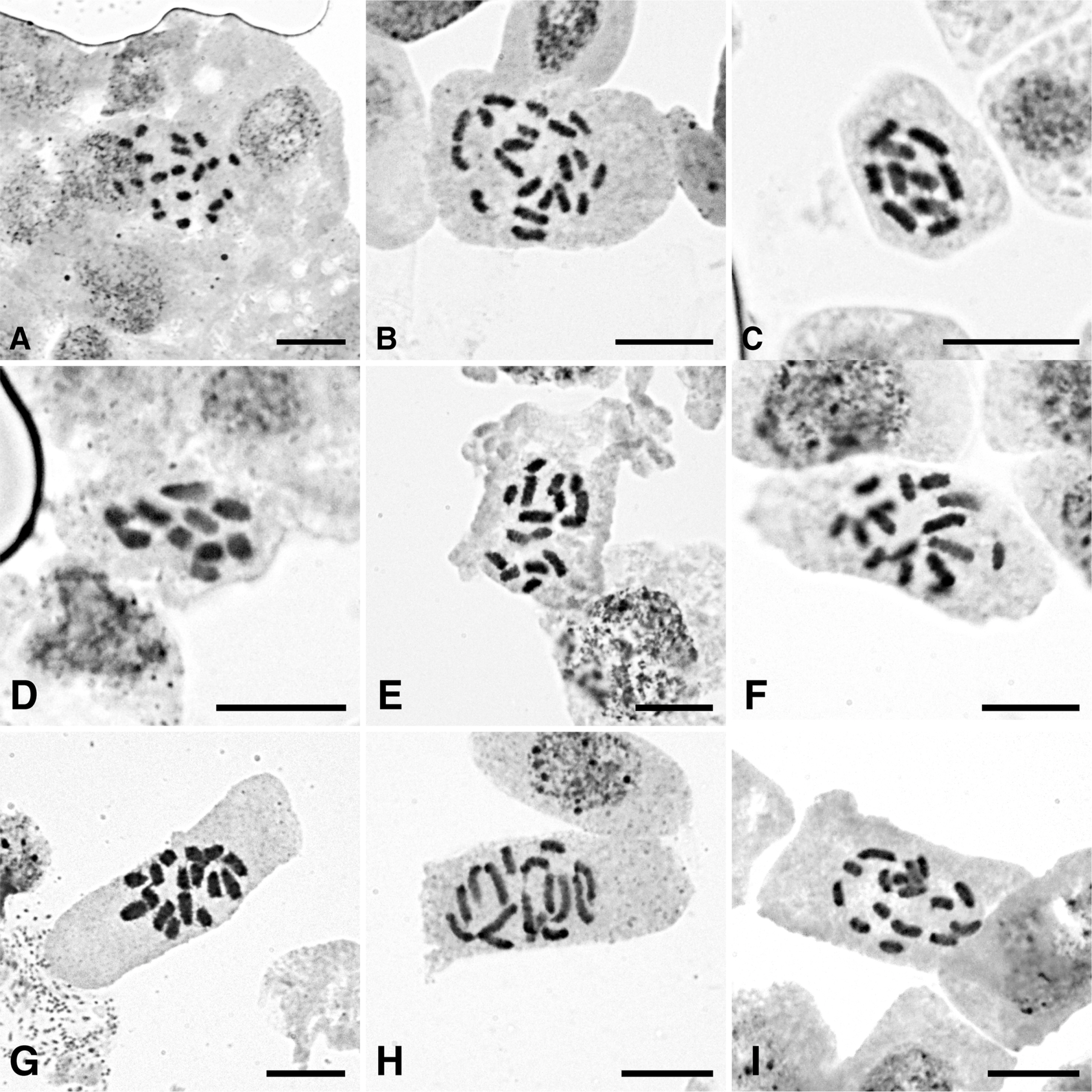
In nine individuals of four Eleocharis taxa, somatic chromosomes were observed (Table 1). In metaphase chromosomes, constrictions were not observed as expected in holocentric chromosomes (Chung and Chung, 2021). The size varies from ca. 1.7 μm to ca. 6 μm. E. attenuata f. laeviseta and E. parvula seem to have more than two types of chromosomes in size, whereas E. ussuriensis and E. valleculosa var. setosa exhibit bimodal karyotypes.
Eleocharis attenuata f. laeviseta H. (2n = 20)
Eleocharis attenuata f. laeviseta from two Jeju Is. populations exhibits the chromosome numbers of 2n = 20 (Fig. 1A, B), which is the first report for the taxon. The size of chromosomes varies along a gradient from ca. 1.7 μm to ca. 4.5 μm. The count is congruent with the chromosome number of E. attenuata f. attenuate (Yano et al., 2004). The taxon occurs in Korea, China, Japan, Russia, Papua New Guinea, and Vietnam, and southern parts of Korean peninsula, Jeollanam-do province and Jeju Is. (Park et al., 2016). Eleocharis attenuata f. laeviseta is distinguished from E. attenuata f. attenuata by depressed style bases and smooth perianth bristles (Hoshino et al., 2011; Park et al., 2016).
Korean name: 참바늘골.
Eleocharis parvula (2n = 10)
We observed 2n = 10 chromosome numbers from two populations of E. parvula in Korea (Fig. 1C, D). The size of chromosomes is ca. 3–4 μm. Previously, two chromosome numbers for the species have been reported from Russian and Japanese populations, 2n = 8 and 2n = 10 (Fedorov, 1969; Yano et al., 2004). The species occurs worldwide in Europe, Siberia, N. Africa, Central, North and South America, and Asia, and habitats are limited to marshes along seacoasts (Dai and Strong, 2010; Hoshino et al., 2011). Recently, Korean populations have been reported from Ganghwa-gun, Incheon Metropolitan City (Ji et al., 2015). The species is characterized by small fusiform tubers, septa-less culms, and confluent style bases with achene apexes, and occurs in salt marshes (Hoshino et al., 2011; Ji et al., 2015).
Korean name: 갯바늘골.
Eleocharis ussuriensis (2n = 16)
Somatic chromosomes of 2n = 16 were observed from the species, confirming previous reports (Fig. 1E, F, Table 1). The size of chromosomes ranges from ca. 2.5 μm to 5 μm in length, grouped in two sizes ca. 2.5–3 μm and ca. 4–5 μm. In the species, Hoshino et al. (1993) observed bimodal karyotypes (small and large) in the species and found satellites in small chromosomes (2.5–3.5 μm in length) from a Chinese population, but they were not found in the Japanese population. Hoshino et al. (2011) also reported the chromosome number of 2n = 20 for the species. However, we are unable to discuss the data because the original article misses voucher species information and chromosome figures. In the chromosomes from Korean populations, satellites were not observed. The species is characterized by long rhizomes and large achenes (1.5–2 μm long), and occurs in Japan, Korea, Sakhalin, Ussuri, and N. E. hina (Hoshino et al., 2011). In Korea, it is found mainly in central areas (Park et al., 2016). The taxon has been treated as E. mamillata var. cyclacarpa in Japan and Korea (Hoshino et al., 2011; Park et al., 2016). However, we used E. ussuriensis based on the principle of publication priority (Dai and Strong, 2010).
Korean name: 물꼬챙이골.
Eleocharis valleculosa var. setosa (2n = 16)
From three populations in Jeju Is., somatic chromosomes of E. valleculosa var. setosa were observed, 2n = 16 (Fig. 1G–I). The size of the chromosomes varies from ca. 3 μm to ca. 6 μm in length, exhibiting bimodal karyotypes, four or five large and the other small ones. The chromosome number agrees with the count made from a Chinese population (Hoshino et al., 1993). E. valleculosa var. valleculosa also has chromosome of 2n = 16 counted from a Japanese population (Yano et al., 2004). The species is characterized by sharp ribs (striate) on culms, oblong spikelets, and four long perianth bristles, growing in shallow waters such as moors and margins of ponds in mountains (Hoshino et al., 2011; Park et al., 2016). It occurs in Korea, China, Japan, and Mongolia (Hohisno et al., 2011). During the research, we examined Eleocharis collections in major herbaria such as Korean National Arboretum and Korea National Institute of Biological Resources herbaria to find E. valleulosa var. valleculosa, which misses perianth bristles. All the herbarium specimens collected and examined were identified as E. valleulosa var. setosa.
Korean name: 까락골.
In four Korean Eleocharis taxa, we observed at least two types of karyotypes, gradient and bimodal. Each karyotype was supported by molecular data although taxon and DNA sampling was limited (Yano et al., 2004). Chromosome numbers of 2n = 10, 16, 20 in Eleocharis suggest polyploidy, aneuploidy, fission and/or fusion events. Because of specialized and reduced morphology and holocentric chromosomes, it is critical to have inclusive taxon and DNA sampling to test traditional classification in a phylogenetic framework. Roalson and Friar (2000) and Yano et al. (2004) attempted to estimate Eleocharis phylogenetic relationships, but the taxon sampling was too limited to apply to the whole genus. Further chromosomal investigation of Korean Eleocharis research should contribute to elucidating phylogenetic relationships of the genus.